螺殼
| 本条目部分链接不符合格式手冊規範。跨語言链接及章節標題等處的链接可能需要清理。(2015年12月11日) |

林場歐洲蝸牛的各向视图:
| 正视图 | 侧视图 |
| 顶视图 | 底视图 |

大型陆生蜗牛(可能是羅馬蝸牛)的截面图:
1 - 脐孔
2 - 壳口处的壳体
3 - 壳口
4 - 轴柱
5 - 缝合线
6 - 螺层
7 - 壳顶
螺殼是指软体动物门腹足纲如蜗牛等动物的外殼,属于一种外骨骼。其作用不仅仅是肌肉的附着物,还有着保护软弱的躯体免受捕食动物或者其它外力的伤害。陆生蜗牛的壳还有遮蔽阳光,免于水分蒸发的作用。螺壳属于贝壳的一种,研究贝壳的学科叫做贝壳学。
绝大多数的螺壳都是成螺旋状,并且其螺旋都是右旋的:从壳顶往下看,旋转方向为順时针。不过也有某些少數类目的螺壳是左旋的:旋轉方向为逆時針。同时存在两种情况的物种,则更为罕见。
螺壳分为多层,其成分通常主要是碳酸钙。这些碳酸钙由软体动物的外套膜所分泌,在沉积到同样由外套膜所分泌的,被称为贝壳素的有机基质上,就变成了坚硬的外壳了。
虽然并非所有腹足纲动物都有壳,但绝大多数都有。而有壳的种群中,绝大部分都只有一片螺旋状的壳体。不过帽贝科中的动物,其成体具有一个简单锥状的壳。
目录
1 旋向
1.1 双旋向物种
2 形成
3 形态
4 度量方式
5 描绘方式
6 详述
7 术语
8 形状分类
9 进化
10 各种形式
11 引用
12 深入阅读
13 外部链接
旋向

两种不同的海螺:左边的是通常为左旋的棱峨螺(已灭绝),而右手边的是通常为右旋的田畦峨螺
由于螺旋状的壳体是非对称性的,因此螺壳具有一个旋向的特征。目前为止一发现的螺壳中,有超过90%都符合右旋则的[1],只有少数种或属中的螺壳是左旋的。而两种情况都有的物种,如雙旋蝸牛属[2],就更为罕见了。分辨左右旋的方法除了前面的顶视法外,还有更简单的方法:将壳尖朝上,螺孔朝向你的脸,此时螺孔在轴线右边的,则是右旋的,反之则为左旋。
腹足纲动物的旋性在胚胎早期的卵裂阶段就已经出现,该性状与NODAL基因有关[3]。
对于通常只有一种旋转方向的物种,在偶然的情况下也会出现旋转方向相反的个体,例如右旋物种中,出现了左旋的个体。贝壳收藏家对这种奇特的壳体是趋之若鹜的。然而螺壳的旋向很容易搞错,因为非专业人员可能会处于图像美观的原因,将图像做了翻转处理。这么一来,一个正常右旋物种的螺壳,就变成了罕见的左旋个体的螺壳了。
双旋向物种
在同一个物种下,同时发现两种旋向的可能并且存在的[4]。尽管右旋物种中,因为基因突变而产生左旋的情况极为罕见,然而人们在陆生蜗牛中确实有这类的记录[4]。而两种旋向在同一个物种中同时大量存在的情况,则更为罕见。到目前为止已确认存在这种情况的,只有个别热带树栖蜗牛的属[4] 。这包括东印度至澳大利亚北部之间的双旋蝸牛属、古巴的丝带蜗牛、海地的糖果蜗牛、夏威夷的小玛瑙螺属中的Partulina科中的某些物种和小瑪瑙螺科的许多物种,以及太平洋小岛上的薄壳蜗牛科薄壳蜗牛属蜗牛等,被证实两种旋向的个体数量均大量同时存在[4]。这些互不相关的群落间都独立出现此类情况,很可能是一个简单的基因突变造成的,尽管这个突变被保留下来的主要原因可能是为了适应树栖生活,而不是为了要改变旋性[4]。在同一个Partula属蜗牛的育囊中,可能会同时出现左旋和右旋的胚胎,尽管通常情况下旋向都应该是一致的。而在双旋蝸牛属中是否有这种遗传特征则没有任何数据[4]。
欧洲的烟管蜗牛科中Alopiinae亚科是一个特例,这是一种习惯在外露石灰岩这种碱性环境上生活的独立群体[4]。它们之中若干个种之所谓被分为不同种的唯一理由,仅仅是因为旋向不同,然而关于左旋和右旋物种是否居住在一起的争取是有争议的[4]。索斯(Soos,1928,pp. 372-385)总结了以前对该问题的讨论,并做出了左旋和右旋是不同的种群,然而也有其他人认为离解答该问题上仍有很长的距离[4]。在秘鲁的烟管蜗牛科Nenia属(皮尔斯布里,1949,pp. 216-217)也被认为是一种双选蜗牛。
德格内尔(Degner,1952)曾经研究过烟管蜗牛科下面一个物种Alinda biplicata罕见的右旋突变基因,这种突变的机制和Radix peregra(缪勒)这种塘螺是相同的——是符合孟德尔定律的一个隐形基因在起的作用,同时任何由杂交所引起的旋向改变会因为某种不明原因而延后一代才被表达[4]。
形成
形态
 螺旋壳体的典型形态。光洁琥珀蜗牛(Zonitoides nitidus,一种陆生右旋蜗牛)的壳的解剖结构。 上: 顶视图,展示螺层和壳顶; 中:侧视图,展示壳的剖面结构; 下: 底视图,展示中心的脐眼。 |  光洁琥珀蜗牛壳实拍(顶视图,侧视图,底视图) |
同一个种的腹足纲动物壳的解剖结构是相当稳定的,除了几个别的情况外,同一科中不同种之间的解剖结构也是相当的稳定的。控制变量包括:
- 每旋转一圈,壳体的生长率。高生长率将会形成大开口形式的螺壳,如鲍鱼。而低生长率则会形成高度螺旋的形态,如锥螺属,或者某些扁蜷科物種;
- 生长曲线的形状,可以近似的看作是壳口的形状。比如说圆形的蝾螺科,狭长形的芋螺科,形状奇异且有延展虹吸沟的骨螺科;
- 生长曲线在轴向上的偏移率。高偏移率形成锥型的螺壳如锥螺属,低偏移率甚至是无偏移率形成扁平的平旋壳,如扁卷螺科,甚至壳口几乎和整个壳体的直径一样,如鲍鱼;
- 不规则性,例如肋、刺、节,以及螺在生长期有规律的改变生长曲线而导致的螺层变化,例如骨螺科的许多属;
- 动物本身成长到成体时的改变,例如女王凤凰螺在变成成体后会有一个外翻的壳唇,而宝螺科则会有一个内翻的壳唇。
这其中的某些因子可以建立数学模型,甚至有计算机程序可以产生十分逼真的螺壳图像。大卫·M·拉普在模拟计算机上曾经进行过这类早期的研究,甚至发现了许多实际上没有被任何腹足纲动物所使用的形态。
在特定的环境中,某些螺壳形状会比其它形状更常见,当然也有许多例外的情况。在高波流能量的环境中,例如布满岩石的潮间带上栖息的海螺,通常壳孔大,壳体高度低。而那些高度螺旋的,甚至带有不规则形状的物种,则通常在精致的水流环境中生存。而在沙滩中穴居的螺如榧螺以及笋螺属等,通常有一个光滑而细长外形,并且没有复杂的不规则外表,这是为了能减少在沙中移动时的阻力。
少数如蛇螺科的腹足纲动物的壳,是与坚固表面如岩石或者其它螺壳粘合在一起,并沿着这些表面生长的。它们的形状会比较奇特,与普通的螺壳差异较大。因此在英文中有另一个名字——虫壳(Worm shell)来形容这一类的螺壳,尽管实际上都是指腹足纲动物的壳。
度量方式

示意14种常用测量方式的顶视图、正视图以及底视图。
在螺壳上进行衡量所使用的常见方式包括:壳的宽、高,以及壳口的宽、高。螺层的数量也是一个常见的度量维度。
这里所说的壳高(或者说长),是指壳的轴线的最大长度,而壳宽则指的是壳体的最大直径。这里的轴是指中轴,壳层围绕这一周线形成螺旋状的螺塔,并通常会在中间形成一个轴柱。上面提到的衡量方式,都是以壳体本身为视角,而不是以壳体内居住动物的视角来描述的。
目前已知最长的壳是澳洲大香螺,这是一种壳体高度可以生长至91厘米的海生蜗牛[5]。
描绘方式

真盘螺的正视图

真盘螺的背视图

真盘螺的底视图

贝氏钟螺(Calliostoma bairdii)活体的顶视图




螺壳可以从多个方向进行展示:
- 正试图:这是常见的展示方式,此时壳口正对读者,壳顶在上方,可以看到壳体的全高。此时如果壳口在纵轴右方,则称为右旋,反之为左旋。
- 背视图:将正视图摆放的螺壳,沿纵轴旋转180°后所成视图。此时壳口背对读者(不可见),壳顶在上方。
- 顶视图:从壳顶往下看,可见壳体全宽。
- 底视图:从壳底往上看,可见壳体全宽。通常该试图仅展示壳体,而不包括生物本身。
详述
壳体是在微小的胚胎时期即开始形成,此时形成的是原胚壳。原胚壳构成螺壳的壳顶尖端,它和壳体的剩余部分通常是有很大差别的。从原胚壳之后开始,壳层会逐渐增大。通常每一段壳层都是圆形或者椭圆形的,然而由于挤压或者其它的原因,也存在许多不同的形状。不同的螺壳,其螺锥或长或短,或广口或细窄,顶角的大小皆不相同。有些螺壳如绮蛳螺的壳层之间并不直接粘连,而另一些螺壳的壳层则会包裹大部分甚至所有旧的壳层,一般来说则介乎于这两者之间。在两条缝合线之间的外层,有的时候会被称为“肩部”。该外露区域的表面可能很光滑,而有的物种则可能会有节、刺等装饰。
最简单的装饰包括随壳体旋转的脊线和螺纹,或者与壳口平行的皱褶和肋条。第一级螺纹是沿着螺旋方向连续的,如果第一级螺纹不在一个平面上,则它们形成的角度就是肩角。第二级螺纹则是第一级之间的纹线,通常尚未成熟的个体不会出现这一级螺纹。某些特殊的物种还会存在第三级螺纹,这些纹线出现在前两级螺纹之间。肋条是横向的突出物,它们之间的间距通常是有规律的,使得每一层的肋条能连成线。在某些特殊的物种里面,它们会在肩角形成之后集中在该肩角上形成节,其余部分都包裹在肩部至体层之间。此后的某个阶段,节可能会转变成刺。这些刺是由之前在壳边上的凹槽在废弃不用之后形成的,这种刺从壳口方向看通常是由开口的。当然,不规则的刺还可以从壳的各种其它部位产生(参见宽角螺)。这些由壳口外唇形成的刺,如果随着壳体螺旋式生长而连成若干排,就会变成纵张肋,这种情况可参见骨螺及许多嵌线螺科的物种。纵张肋还有另一种形成方式,即在静止期可口外唇向外扩张。这种纵张肋具体分为两种类型,一种是如绮蛳螺般形成一个较薄的平面,另一种是如竖琴螺属般壳口外唇向外翻卷形成一圆条。某些物种如骨螺或者蟹守螺属,它们会在特定位置扩大螺孔,尽管这样形成了若干排的“节”,但这并不会被归类为纵张肋。
壳口的形状可能很简单,也可能千奇百怪。壳口的外唇和内唇很容易被人识别。这两者通常是相互融合的,但也可能在壳底部有一个切口,甚至如纺锤螺属那般向外突出并形成或长或短的虹管。在某些类型的螺壳上会出现向上或者说向后方开口的凹槽,这种形式主要出现在较老的品种身上。这种情形将会导致在缝合线边上形成一个脊,或者托架状的装饰(如棒螺)。某些物种如翁戎螺科、神螺科等,在外唇上的切口会一直延展下去,进而形成一条很长的裂缝。而这些裂缝的不断闭合,会形成明显案的裂缝带。某些情况下这些废弃的裂缝会形成一个孔(如钥孔䗩科),或者因周期性产生裂缝而形成一系列的孔(如鲍螺属)。这种外表面的缺陷通常可以指出其生长线。
在外唇内侧有时候能发现成为旋文的脊或者皱褶,有些还比较坚硬如牙齿一般(如蜒螺)。与之类似的脊或者皱褶在内唇里面贴近壳轴的壳壁上更常见,称之为轴旋褶。这种皱褶通常和壳轴相垂直或者斜交,只有极少数品种是在壳口处轻易可见,大部分都会隐藏在远离壳口的内部因而在完整的体中是不可直接看见的,除非将其打碎。
当壳轴是空心时(壳体未紧密结合,形似在中间钻了一个狭长的小洞),在低端所形成的开口被称为脐孔。脐孔的形式和大小也是多种多样的,甚至可能完全或者部分的被内唇的延长物所覆盖(如玉螺属)。
绝大多数现存的螺壳外面都会有覆盖物,例如光滑的角质层,或者有披毛的表皮层,再或者外壳膜,有的时候这些覆盖物还会遮盖壳体上的绚丽色彩。这些覆盖物以及壳体上的色彩通常很难在化石中得以保存,因此很难断定古代的螺壳是否也有如此结构。
腹足纲动物近壳口处称为前端,反之螺塔塔尖处称为后端。大部分的插图作者通常将壳尖画成朝上,但是法国作者则通常画成朝下。壳口通常会有一角质或者碳酸钙质的螺盖,不同的物种间形态各异。螺盖通常产生并附着于动物腹足背面上方。[6]
术语

图为一个形状不寻常的散大蜗牛(Helix aspersa)的壳
用于描述螺壳各部分的术语包括:
殼口: 螺壳的开口;
殼頂: 壳顶端若干极小的壳层,通常包含胚壳;
体层: 壳体中最大的一层壳层,腹足纲动物的内脏部即藏于此中;
轴柱: 壳体绕轴旋转生长时,在轴上形成的柱体;
壳盖: 壳口处的薄片,用于盖住壳口;
壳胝: 某些腹足纲物种的壳口轴壁上会延伸出来的一个壳体结构,起作用类似于底座;
外壳膜: 许多腹足纲物种的壳体外表会覆盖一层生物层的“薄皮肤”;
围口: 壳口一圈的部分;
胚壳: 幼体时的壳,通常长为成体之后仍会保留在壳顶处;
装饰: 壳层表面凹凸如装饰的结构;
纹线: 壳体外表面上的纹线,属于其中一种装饰;
口褶: 壳口轴壁上的皱褶,也是一种装饰;
虹管: 某些腹足纲物种的壳口轴壁会延伸成细长中空壳体结构;
螺塔: 体层以上的壳体结构;
缝合线: 在大多数腹足纲动物壳体上,各壳层之间会连接在一起形成一条沟槽;
脐孔: 由于壳体通常会延中轴旋转生长,因此在壳底面中心会有一个深凹的结构称为脐孔,在壳体内与之对应的即为轴柱;
纵张肋: 某些腹足纲动物在生长静止期时,壳口会变厚,于是形成一个纵向的突出肋条状结构,使壳体呈现若干条“曲张静脉”,即为纵张肋;
壳层: 壳体每旋转一圈,即形成一层壳层。
形状分类
形状的分类方式取决于关注哪些方面,例如,根据高宽比的不同,可以将壳的形状分为三组[7]:
- 长锥形——高度远超宽度;
- 球状或者锥形——宽度和高度基本相同;
- 扁平形——宽度远超高度。

长锥形的Bulgarica denticulata

球形的Sphincterochila candidissima

扁平形的坎佩尔田螺

en:Turritella communis



以下是详细的形状分类:[8][9]

帽形

耳形

蜒螺形

平螺旋形

盘螺形

马蹄螺形(笋型)

圆润锥形*

锥形*

平滑锥型(开酒器形)

塔顶形

纺锤形——现存蜗牛中壳体最大的澳洲大香螺

马球杆形

蛋形

不规则形











* 某些文献中提到的梨形指的是圆润锥形及锥形
进化
螺壳形式的多样化,可能是由各种不同的原因造成的。引起这种进化的原因包括物理稳定性[10],抵御捕食者[11],性选择[12],以及气候选择性[13][14]等。
某些腹足纲动物的壳在进化的过程中退化甚至消失了,例如所有的蛞蝓,半蛞蝓以及许多其它物种。有时候这种壳体的退化,和捕食性的进食方式有关。某些物种在进化过程中,壳体的螺旋性甚至消失了[15] 。根据道罗法则,失去的螺旋特性不会再次重新获得。然而,在帆螺科中 确实有一些属通过改变发育时间(异时性),在原有条件下没有螺旋的帽状贝克的基础上,重新获得了带有螺旋的壳(重进化)[15]。
各种形式

有许多壳层的欧洲锥螺的壳

欧洲锥螺壳的X射线图像

海生贝蜗牛埃及宝螺的壳

埃及宝螺壳的X射线图像




栗色鹑螺壳的X射线图像

某种法螺属壳的X射线图像

维那斯骨螺壳的X射线图像
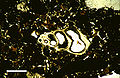
一片来自圣萨尔瓦多岛的,年代为全新世的泻湖沉积物的薄切片,在平面偏振光显微镜下可见微小腹足纲动物的壳,放大比例为5万倍。





带有壳盖的淡水蜗牛大羊角螺的壳是平面螺旋的。

可食用有肺类陆生罗马蜗牛的壳和生物体。

海生中华蝾螺的壳,图中上排展示了壳盖。

海生帽贝的壳,可能是一个欧洲帽贝物种。




引用
本文使用了公众领域中的文字[4][6],以及符合CC-BY-2.0版权声明的文字[14]。
^ Schilthuizen, M.; Davison, A. The convoluted evolution of snail chirality. Naturwissenschaften(德国《自然科学》杂志). 2005, 92 (11): 504–515. doi:10.1007/s00114-005-0045-2 (英语).
^ Amphidromus perversus (Linnaeus, 1758)
^ Myers P. Z. (13 April 2009) "Snails have nodal!". The Panda's Thumb, accessed 3 May 2009.
^ 4.004.014.024.034.044.054.064.074.084.094.10 [[
File:PD-icon.svg|15px]] Laidlaw F. F. & Solem A. (1961). "The land snail genus Amphidromus: a synoptic catalogue". Fieldiana Zoology 41(4): 505-720.
^ Wells F. E., Walker D. I. & Jones D. S. (eds.) (2003) "Food of giants – field observations on the diet ofSyrinx aruanus(Linnaeus, 1758) (Turbinellidae) the largest living gastropod". The Marine Flora and Fauna of Dampier, Western Australia. Western Australian Museum, Perth.
^ 6.06.1 Grabau A. W. & Shimer H. W. (1909) North American Index Fossils Invertebrates. Volume I.. A. G. Seiler & Company, New York. pages page 582-584.
^ Falkner G., Obrdlík P., Castella E. & Speight M. C. D. (2001). Shelled Gastropoda of Western Europe. München: Friedrich-Held-Gesellschaft, 267 pp.
^ Hershler R. & Ponder W. F.(1998). "A Review of Morphological Characters of Hydrobioid Snails". Smithsonian Contributions to Zoology 600: 1-55. [1].
^ Dance P. S. (). Shells.
^ Britton J. C (1995) "The relationship between position on shore and shell ornamentation in 2 size-dependent morphotypes of Littorina striata, with an estimate of evaporative water-loss in these morphotypes and in Melarhaphe neritoides". Hydrobiologia 309: 129-142. abstract.
^ Wilson A. B., Glaubrecht M. & Meyer A. (March 2004) "Ancient lakes as evolutionary reservoirs: evidence from the thalassoid gastropods of Lake Tanganyika". Proceeings of Royal Society London Series B - Biological Sciences 271: 529-536. doi:10.1098/rspb.2003.2624.
^ Schilthuizen M. (5 June 2003) "Sexual selection on land snail shell ornamentation: a hypothesis that may explain shell diversity".BMC Evolutionary Biology 3: 13. doi:10.1186/1471-2148-3-13.
^ Goodfriend G. A. (1986) "Variation in land-snail shell form and size and its causes – a Review". Systematic Zoology 35: 204-223.
^ 14.014.1 Pfenninger M., Hrabáková M., Steinke D. & Dèpraz A. (4 November 2005) "Why do snails have hairs? A Bayesian inference of character evolution". BMC Evolutionary Biology 5: 59. doi:10.1186/1471-2148-5-59
^ 15.015.1 Collin R. & Cipriani R. (22 December 2003) "Dollo’s law and the re-evolution of shell coiling". Proceedings of the Royal Society B 270(1533): 2551-2555.doi:10.1098/rspb.2003.2517.
深入阅读
- 关于旋向
- van Batenburg1 F. H. D. & Gittenberger E. (1996). "Ease of fixation of a change in coiling: computer experiments on chirality in snails". Heredity 76: 278-286. doi:10.1038/hdy.1996.41.
- Wandelt J. & Nagy L. M. (24 August 2004) "Left-Right Asymmetry: More Than One Way to Coil a Shell". Current Biology 14(16): R654-R656. doi:10.1016/j.cub.2004.08.010
外部链接
Gastropods by J. H. Leal - Information on some gastropods of the tropical Western Atlantic, specifically the Caribbean Sea, with relevance to the fisheries in that region- Radiocarbon Dating of Gastropod Shells
- Nair K. K. & Muthe P. T. (18 November 1961) "Effect of Ribonuclease on Shell Regeneration in Ariophanta sp.". Nature 192: 674-675. doi:10.1038/192674b0.
(西班牙文) Antonio Ruiz Ruiz, Ángel Cárcaba Pozo, Ana I. Porras Crevillen & José R. Arrébola Burgos Caracoles Terrestres de Andalúcia. Guía y manual de identificación. 303 pp., ISBN 84-935194-2-1. (from website)


