Competitive exclusion principle

1: A smaller (yellow) species of bird forages across the whole tree.
2: A larger (red) species competes for resources.
3: Red dominates in the middle for the more abundant resources. Yellow adapts to a new niche restricted to the top and bottom of the tree, avoiding competition.
In ecology, the competitive exclusion principle,[1] sometimes referred to as Gause's law,[2] is a proposition named for Georgy Gause that two species competing for the same limiting resource cannot coexist at constant population values. When one species has even the slightest advantage over another, the one with the advantage will dominate in the long term. This leads either to the extinction of the weaker competitor or to an evolutionary or behavioral shift toward a different ecological niche. The principle has been paraphrased in the maxim "complete competitors cannot coexist".[1]
Contents
1 History
2 Experimental basis
3 Prediction
4 Paradoxical traits
5 Redefinition
6 Application to humans
7 See also
8 References
History
The competitive exclusion principle is classically attributed to Georgii Gause,[3] although he actually never formulated it.[1] The principle is already present in Darwin's theory of natural selection.[2][4]
Throughout its history, the status of the principle has oscillated between a priori ('two species coexisting must have different niches') and experimental truth ('we find that species coexisting do have different niches').[2]
Experimental basis

Paramecium aurelia and Paramecium caudatum grow well individually, but when they compete for the same resources, P. aurelia outcompetes P. caudatum.
Based on field observations, Joseph Grinnell formulated the principle of competitive exclusion in 1904: "Two species of approximately the same food habits are not likely to remain long evenly balanced in numbers in the same region. One will crowd out the other".[5]Georgy Gause formulated the law of competitive exclusion based on laboratory competition experiments using two species of Paramecium, P. aurelia and P. caudatum. The conditions were to add fresh water every day and input a constant flow of food. Although P. caudatum initially dominated, P. aurelia recovered and subsequently drove P. caudatum extinct via exploitative resource competition. However, Gause was able to let the P. caudatum survive by differing the environmental parameters (food, water). Thus, Gause's law is valid only if the ecological factors are constant.
Gause also studied competition between two species of yeast, finding that Saccharomyces cerevisiae consistently outcompeted Schizosaccharomyces kefir[clarification needed] by producing a higher concentration of ethyl alcohol.[6]
Prediction
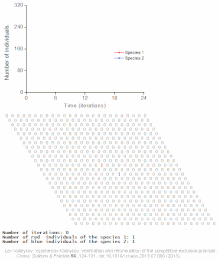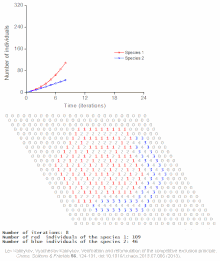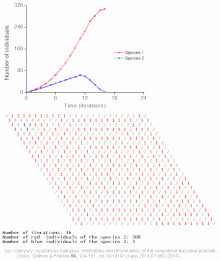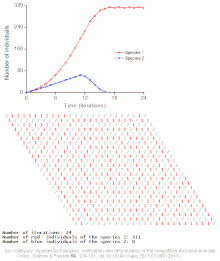
Cellular automaton model of interspecific competition for a single limited resource
Competitive exclusion is predicted by mathematical and theoretical models such as the Lotka-Volterra models of competition. However, for poorly understood reasons, competitive exclusion is rarely observed in natural ecosystems, and many biological communities appear to violate Gause's law. The best-known example is the so-called "paradox of the plankton".[7] All plankton species live on a very limited number of resources, primarily solar energy and minerals dissolved in the water. According to the competitive exclusion principle, only a small number of plankton species should be able to coexist on these resources. Nevertheless, large numbers of plankton species coexist within small regions of open sea.
Some communities that appear to uphold the competitive exclusion principle are MacArthur's warblers[8] and Darwin's finches,[9] though the latter still overlap ecologically very strongly, being only affected negatively by competition under extreme conditions.[10]
Paradoxical traits
A partial solution to the paradox lies in raising the dimensionality of the system. Spatial heterogeneity, trophic interactions, multiple resource competition, competition-colonization trade-offs, and lag may prevent exclusion (ignoring stochastic extinction over longer time-frames). However, such systems tend to be analytically intractable. In addition, many can, in theory, support an unlimited number of species. A new paradox is created: Most well-known models that allow for stable coexistence allow for unlimited number of species to coexist, yet, in nature, any community contains just a handful of species.
Redefinition
Recent studies addressing some of the assumptions made for the models predicting competitive exclusion have shown these assumptions need to be reconsidered. For example, a slight modification of the assumption of how growth and body size are related leads to a different conclusion, namely that, for a given ecosystem, a certain range of species may coexist while others become outcompeted.[11][12]
One of the primary ways niche-sharing species can coexist is the competition-colonization trade-off. In other words, species that are better competitors will be specialists, whereas species that are better colonizers are more likely to be generalists. Host-parasite models are effective ways of examining this relationship, using host transfer events. There seem to be two places where the ability to colonize differs in ecologically closely related species. In feather lice, Bush and Clayton[13] provided some verification of this by showing two closely related genera of lice are nearly equal in their ability to colonize new host pigeons once transferred. Harbison[14] continued this line of thought by investigating whether the two genera differed in their ability to transfer. This research focused primarily on determining how colonization occurs and why wing lice are better colonizers than body lice. Vertical transfer is the most common occurrence, between parent and offspring, and is much-studied and well understood. Horizontal transfer is difficult to measure, but in lice seems to occur via phoresis or the "hitchhiking" of one species on another. Harbison found that body lice are less adept at phoresis and excel competitively, whereas wing lice excel in colonization.
Application to humans
Evidence showing that the competitive exclusion principle operates in human groups has been reviewed and integrated into regality theory to explain warlike and peaceful societies[15]. For example, hunter-gatherer groups surrounded by other hunter-gatherer groups in the same ecological niche will fight, at least occasionally, while hunter-gatherer groups surrounded by groups with a different means of subsistence can coexist peacefully[15].
See also
- Limiting factor
- Limiting similarity
- Paradox of the plankton
References
^ abc Garrett Hardin (1960). "The competitive exclusion principle" (PDF). Science. 131 (3409): 1292–1297. doi:10.1126/science.131.3409.1292. PMID 14399717..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ abc Pocheville, Arnaud (2015). "The Ecological Niche: History and Recent Controversies". In Heams, Thomas; Huneman, Philippe; Lecointre, Guillaume; et al. Handbook of Evolutionary Thinking in the Sciences. Dordrecht: Springer. pp. 547–586. ISBN 978-94-017-9014-7.
^ Gause, Georgii Frantsevich (1934). The Struggle For Existence (1st ed.). Baltimore: Williams & Wilkins.
^ Darwin, Charles (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (1st ed.). London: John Murray. ISBN 1-4353-9386-4.
^ Grinnell, J. (1904). "The Origin and Distribution of the Chestnut-Backed Chickadee". The Auk. American Ornithologists' Union. 21 (3): 364–382. doi:10.2307/4070199. JSTOR 4070199.
^ Gause, G.F. (1932). "Experimental studies on the struggle for existence: 1. Mixed population of two species of yeast" (PDF). Journal of Experimental Biology. 9: 389–402.
^ Hutchinson, George Evelyn (1961). "The paradox of the plankton". American Naturalist. 95: 137–145. doi:10.1086/282171.
^ MacArthur, R.H. (1958). "Population ecology of some warblers of northeastern coniferous forests". Ecology. 39: 599–619. doi:10.2307/1931600.
^ Lack, D.L. (1945). "The Galapagos finches (Geospizinae); a study in variation". Occasional Papers of the California Academy of Sciences. 21: 36–49.
^ De León, LF; Podos, J; Gardezi, T; Herrel, A; Hendry, AP (Jun 2014). "Darwin's finches and their diet niches: the sympatric coexistence of imperfect generalists". J Evol Biol. 27 (6): 1093–104. doi:10.1111/jeb.12383.
^ Rastetter, E.B.; Ågren, G.I. (2002). "Changes in individual allometry can lead to coexistence without niche separation". Ecosystems. 5: 789–801. doi:10.1007/s10021-002-0188-3.
^ Moll, J.D.; Brown, J.S. (2008). "Competition and Coexistence with Multiple Life-History Stages". American Naturalist. 171: 839–843. doi:10.1086/587517.
^ Clayton, D.H.; Bush, S.E. (2006). "The role of body size in host specificity: Reciprocal transfer experiments with feather lice". Evolution. 60: 2158–2167. doi:10.1111/j.0014-3820.2006.tb01853.x.
^ Harbison, C.W. (2008). "Comparative transmission dynamics of competing parasite species". Ecology. 89: 3186–3194. doi:10.1890/07-1745.1.
^ ab Fog, Agner (2017). Warlike and Peaceful Societies: The Interaction of Genes and Culture. Open Book Publishers. doi:10.11647/OBP.0128. ISBN 978-1-78374-403-9.
Biological rules | ||
|---|---|---|
| Rules |
|  |
| Related |
| |


