恐龙
| 恐龍 化石时期:231.4–0Ma PreЄ Є O S D C P T J K Pg N 三疊紀晚期–現今 (包含鳥類) | |
|---|---|
 | |
| 恐龙骨架组图,由左上顺时针:小盗龙、迷惑龙、剑龙、三角龙、埃德蒙顿龙、加斯顿龙 | |
科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 爬行纲 Reptilia |
| 亚纲: | 双孔亚纲 Diapsida |
| 演化支: | 新双弓类 Neodiapsida |
| 演化支: | 蜥类 Sauria |
| 下纲: | 主龙形下纲 Archosauromorpha |
| 演化支: | 主龙类 Archosauria |
| 演化支: | 鸟跖类 Avemetatarsalia |
| 演化支: | 鸟颈类主龙 Ornithodira |
| 演化支: | 恐龙形态类 Dinosauromorpha |
| 演化支: | 恐龙形类 Dinosauriformes |
| 演化支: | 恐龙总目 Dinosauria Owen, 1842 |
目 | |
| |
| 古生物 |
| 古生物學 |
|---|
 |
化石 化石 · 遗迹化石 · 指準化石 · 化石点列表 · Lagerstätte fossil beds · List of transitional fossils · List of human evolution fossils |
自然歷史 生物地理學 · 生物集群滅絕 · Geochronology · 地質年代 · Geologic record · History of life · 生命起源 · 生命演化歷程 · 過渡化石 |
器官与演化 Avian flight · Cells · 多細胞生物 · Eyes · Flagella · Hair · 哺乳動物聽小骨進化 · Mosaic evolution · Nervous systems · 有性生殖的演化 |
不同類群的演化 鸟的进化 · Butterflies · Cephalopods · Cetaceans · 恐龍 · Fishes · Fungi · 人類演化 · Insects · Mammals · Molluscs · Plants · Reptiles · Sea cows · Spiders · 四足類 |
進化 Introduction to evolution · 共同起源 · 系統發生樹 · 支序分類學 · 生物分類法 |
古生物史 古生物史 · Timeline of paleontology |
分支 Biostratigraphy · Ichnology · Invertebrate paleontology · 微體古生物學 · Molecular paleontology · 木化石 · Paleobiology · 古植物學 · 古生態學 · Paleogenetics · Paleolimnology · Paleopedology · Paleotempestology · 古動物學 · 孢粉學 · Sclerochronology · Taphonomy · Vertebrate paleontology |
古生物 古生物學 |

簡略版的恐龍演化樹
恐龙是恐龍總目(學名:Dinosauria)的通称,包括了已灭绝的非鳥型恐龙(英语:Non-avian dinosaurs,又稱非鳥恐龍)及现生的鸟类,是出現於中生代多樣化優勢陸棲脊椎動物,曾支配全球陸地生态系统超過1亿6千万年之久。恐龙最早出现在2亿3千万年前的三疊紀,大部份於约6千5百万年前的白垩纪晚期所发生的白垩纪末滅絕事件中絕滅,僅有倖存「鸟型恐龙」(英语:Avian dinosaurs),即现今的鳥类存活下来[1][2]。
自从19世纪的工业革命早期,第一批恐龙化石被科學方法鑑定後,重建的恐龙骨架因为其体型极其巨大或小巧、构造奇妙,已成為全球各地博物馆的主要展覽品,這古代生物開始為世人所知。在20世紀前半期,随着电影工业在美国兴起,大眾媒體都視恐龍為行動緩慢、慵懶的冷血動物。但是1970年代開始的恐龍文藝復興,提出恐龍也許是群活躍的溫血動物,並可能有社會行為。近期發現的眾多恐龍與鳥類之間關係的證據,支持了恐龍溫血動物的假設。恐龙已是大眾文化的一部分,无论儿童或者成年人均对恐龙有很高的兴致。恐龙往往是热门书籍與电影的题材,如:《侏罗纪公园》系列电影,各类媒体也常報導恐龙的科学研究進展與新發現。
1861年,考古学家發現的身为鸟类的始祖鳥化石、却與身为恐龙的美頜龍化石極度相似,差別只在於始祖鳥化石有著羽毛痕跡,這顯示恐龍與鳥類可能是近親。1970年代以來,許多研究指出现代鸟类極可能是蜥臀目兽脚亚目虚骨龙类近鳥型恐龙的直系後代[3][4]。1990年代中後期,古生物學家開始在中國發現大量的化石證據,中華龍鳥、尾羽龍、北票龍等有直接羽毛證據的有羽毛恐龍相繼被發現,為恐龍與鳥類間的演化關係提供了愈漸明確的線索,而如學界普遍都視鳥類為恐龙的直系后代[5],而甚至有少數科學家主張牠們應該分類於同一綱之內[6];於此同時,因为孔子鸟等鸟类和恐龙的中间物种也相继被发现、填补了原本化石材料上的空白,更加确定了鸟类和恐龙之间的演化关系,导致鸟类从“恐龙的后代”進一步被認定为“唯一幸存发展至今的恐龙总目直系”。
許多史前爬行動物常被一般大眾非正式地認定是恐龙,例如:翼龍、魚龍、蛇頸龍、滄龍、盤龍類(異齒龍與基龍)等,但从嚴謹的科学角度来看这些都不是恐龍,反倒是雞、鴨、孔雀才是真正的恐龍。翼龍和恐龍是這幾個物種裡面關係最近的近親,都屬於鳥頸類;恐龍和翼龍是鱷魚、蛇頸龍的遠親,鱷魚所屬的鱷目、和蛇頸龍所屬的鰭龍超目,和恐龍翼龍所屬的鳥頸類同屬於主龍類;恐龍、翼龍、鱷魚、蛇頸龍所屬的主龍類和滄龍是關係較遠的物種,他們和滄龍所屬的有鱗目同屬蜥類;最後,恐龍、翼龍、鱷魚、蛇頸龍、滄龍他們和魚龍是關係很遠的物種,唯一的聯繫是都屬於蜥形綱的一分子。
目录
1 概述
1.1 命名
1.2 鑑定特徵
1.3 種系發生學定義
2 演化與分類
2.1 恐龍的起源
2.2 科学分类
2.3 演化與地理分佈
2.4 白堊紀晚期的多樣性降低?
3 古生物學
3.1 體型
3.1.1 最大和最小的恐龙
3.2 行為
3.3 生理特徵
3.4 軟組織與DNA
4 與鳥類的關係
4.1 有羽毛恐龍
4.2 骨骸
4.3 軟組織
4.4 繁殖
4.5 行為證據
5 灭绝理论
5.1 撞击事件
5.2 火山爆發
5.3 生態系統變遷
5.4 古新世倖存者?
6 发掘历史
6.1 恐龍文藝復興
7 大眾文化
7.1 衍生用語
8 宗教观点
9 相关条目
10 参考文献
11 延伸閱讀
12 外部链接
概述

早期的斑龍骨架圖繪,斑龍是最早被正式命名的恐龍,也是「恐龍」一詞出現的開端
恐龍(包含鳥類)是群生存於陸地上的主龍類爬行動物,四肢直立於身體之下,而非往兩旁撐開,牠們出現於晚三疊紀卡尼階,各種恐龍快速地演化出不同的特化特徵,並發展出不同的體型大小,佔據著不同的生態位,並持續生存到晚白堊紀馬斯垂克階[7]。有許多型態的爬行動物與恐龍生存在同一時期,例如:魚龍目、滄龍科、蛇頸龍目、翼龍目、以及盤龍目,許多大眾媒體常將牠們也歸類於恐龍,但牠們在科學分類上並不屬於恐龍。魚龍類、滄龍類、蛇頸龍類都是海生爬行動物,並不生存於陸地上,也不屬於主龍類;翼龍類屬於主龍類,是在三疊紀時期與恐龍分開演化,但並非陸棲動物:盤龍類生存於二疊紀,親緣關係較接近於哺乳類[8]。
恐龍是中生代的優勢陸棲脊椎動物,尤其是在侏羅紀與白堊紀。在恐龍佔優勢的侏羅紀與白堊紀期間,幾乎所有身長超過1公尺的陸地動物皆為恐龍,同時代的其他動物因此受到體型發展與生態位上的限制。舉哺乳類為例,牠們的體型多接近現代的囓齒動物,不超過貓的大小,食性為肉食性,以小型動物為生[9]。生存在白堊紀的三椎齒獸目的巨型爬獸(Repenomamus giganticus)是個少數例外,巨型爬獸體重12到14公斤,已知會以幼年鸚鵡嘴龍等小型恐龍為食[10]。
恐龙家族极为庞大,因为不仅仅包含了已灭绝的恐龙、还包含了现代的鸟类,所以其物种多样性极其丰富。截止到2006年的学术研究,已確定有超過500个屬;根據估計,化石記錄中曾出現的屬总数約为1850個,當中有75%已被發現化石[11]。一個早期的研究推測恐龍有將近3,400個屬,但大部分無法在化石記錄中保存下來[12]。截至2008年9月17日為止,恐龍記有1,047個種[13]。恐龍有植食性動物,也有肉食性、雜食性動物。有些恐龍以双足行走,或四足行走,或如砂龙和禽龙可以在双足和四足间自由转换。許多恐龍的身上具有鱗甲,或是頭部長有角或頭冠。儘管恐龍以其巨大體型而著稱,但許多恐龍的體型只有人類大小,甚至更小。目前已在全球各大洲發現恐龍化石,包含南極洲在內[14]。无论体型大小,恐龙对陆地生活的适应性堪称卓越,但牠們無法佔據海生以及飛行動物的生態位。
命名

「恐龍」一詞的命名者,理查·歐文
歷史上可追溯到最早的恐龍化石明確紀錄是在1677年,由當時英國牛津大學的博物學家羅伯特‧普勞特發現的恐龍化石,這批化石後來於1824年由英國地質學家威廉·布克蘭命名為斑龍。[15]
恐龙(Dinosauria)是在1842年由英国古生物学家理查·欧文正式提出,歐文把恐龍總目定义为蜥形纲(Sauropsida)的一个独特部族或亚目,包含當時在英國與世界各地發現的恐龍[16]。Dinosauria這名詞衍化自古希腊文,δεινός/deinos意为「恐怖的」或「极其巨大的」,σαύρα/sauros意为「蜥蜴」[17]。欧文在命名時,考量到牠們的牙齒、利爪、巨大体型、以及其他令人印象深刻的恐怖特徵,因此命名牠們為「恐怖的蜥蜴」[18]。中文的恐龙一词,則源自日本對Dinosauria一詞的翻译「恐龍(恐竜)」,最初由中國地質學家章鴻釗提出[19],后来在中文地区广泛使用。
鑑定特徵
近年來的新發現化石,使得科學家們越來越難列出一致性的恐龍鑑定特徵。恐龍的骨骼具有一些演化適應,可使牠們與原始的主龍類祖先辨別出來。在後期的恐龍中,這些特徵經過更多的演化改變。
恐龍的共有衍徵包含:
肱骨有低矮的三角嵴(Deltopectoral crest,讓胸鎖三角肌肉附著的部分),長度約是肱骨的1/3到1/2。
腸骨後部有個突出區塊。
脛骨末端邊緣寬廣,有個往後的凸緣。
距骨有個明顯上突,與脛骨契合[20]。
除上述幾個特徵以外,大部分恐龍還有一些共同特徵,但是因為出現在其他主龍類,或者不存在於早期的恐龍身上,因此不列為恐龍的共有衍徵。舉例而言,恐龍具有兩對顳顬孔,這是雙孔動物的共同特徵,恐龍還具有眶前孔與下頜孔,這些則是主龍類的共同特徵[21]。另有數個特徵,早在恐龍之前已經出現,消失於最早期的恐龍,而在不同的恐龍生物群獨自演化出現,包含:延長的肩胛骨、至少三個癒合的薦椎(其他主龍類也有三個癒合薦椎,艾雷拉龍只有兩個癒合薦椎)[20]、開放式有孔的髖臼(農神龍的髖臼閉合)[22]。恐龍的鑑定特徵難以確認,部分原因則是晚三疊紀的早期恐龍與同時期其他主龍類的化石不多,兩者卻有許多相似處,使得某些主龍類曾被誤認為是恐龍[23]。

三種不同的臀窩結構與後肢直立方式,從左到右分別為往兩側延展、直立(哺乳類與恐龍)、柱狀直立方式(勞氏鱷目)
恐龍具有直立的步態,類似大部分的現代哺乳類,而大部分其他爬行動物則是四肢往兩側延展的步態[24]。恐龍的臀窩朝向兩側,股骨的第四粗隆部往內側,兩者契合,產生直立的步態[25]。直立的步態可讓恐龍快速運動時,可以同時呼吸,使牠們成為活躍的動物,較採取兩側延展步態的動物更具優勢[26]。直立步態可以降低四肢彎曲時所承受的壓力,有助於恐龍發展出巨大的體型[27]。某些非恐龍的主龍類也獨自發展出直立的步態,例如勞氏鱷目,但牠們的臀窩朝下,股骨往上嵌入臀窩,形成不同於恐龍與哺乳類的「柱狀直立方式」[27]。
種系發生學定義

劍龍骨架,位於芝加哥菲爾德博物館
根據種系發生學的定義,恐龍包含:三角龍與現代鳥類的最近共同祖先,以及其最近共同祖先的所有後代[28]。在過去,恐龍曾被定義為:斑龍、禽龍與林龍的最近共同祖先,以及其最近共同祖先的所有後代,以上三種恐龍即為理查·歐文最初用來定義恐龍範圍的三個屬[29]。兩個定義的範圍相近,都包含獸腳亞目、蜥腳形亞目、劍龍下目、甲龍下目、角龍下目、厚頭龍下目、以及鳥腳下目等演化支。早在近年種系發生學研究盛行前,上述分類單元皆已在科學文獻中出現、使用,並持續到今日。
現今古生物學界普遍同意鳥類是恐龍的直系後代。根據嚴格的親緣分支分類法定義,一個天然的演化支應包含單一共同祖先的所有後代。若以此方法來看,所有鳥類都為恐龍,而恐龍並沒有滅亡。大部分古生物學家將鳥類歸類於恐龍-蜥臀目-獸腳亞目-虛骨龍類-手盜龍類[30]。
根據親緣分支分類法,鳥類都是恐龍。但對於一般大眾與媒體而言,恐龍並不包含鳥類。因此出現了「非鳥類恐龍」一詞,以指名不包含鳥類的恐龍,並符合林奈式分類法中的恐龍範圍。林奈氏分類法允許並系群的存在。
演化與分類
恐龍的起源

馬拉鱷龍,是種三疊紀的鳥頸類主龍
恐龍在過去被認為是複系群,包含數個毫不相干的演化支,並非來自於共同的最近祖先,因為身處類似的生態環境中,而個別演化出類似的形態與特徵[31][32][33]。自從70年代的恐龍文藝復興以來,恐龍已被認為是單系群,具有最近的共同祖先,是個天然演化支[34][20]。
恐龍與牠們的主龍類祖先在三疊紀中到晚期分開演化,接近2億3000萬年前,大約是二疊紀-三疊紀滅絕事件後約2000萬年,該次滅絕事件消滅了接近95%的地球生物物種[35][36]。對於早期恐龍始盜龍發現地層的放射線定年法檢驗,證實始盜龍生存於中三疊紀。古生物學家認為始盜龍的外表類似所有恐龍的共同祖先[37];如果屬實,則顯示最早的恐龍應該是種小型、二足掠食動物[38]。在阿根廷的中三疊紀地層所發現的原始、類似恐龍的鳥頸類主龍,例如:馬拉鱷龍與兔蜥,也支持這個說法;已出土的化石證據顯示這群動物都為小型、二足掠食動物。
恐龍剛出現時,陸地上有多種不同的動物,例如堅蜥目、勞氏鱷目、鳥鱷科、喙頭龍目等原始主龍類,以及二齒到底獸下目與犬齒獸亞目等獸孔目動物。這些動物大部分在三疊紀後期的兩次滅絕事件中滅亡。第一次滅絕事件發生於卡尼階與諾利階交接處,約2億1500萬年前,二齒獸類與許多原始主龍形下綱動物在此事件中滅亡,例如原蜥形目與喙頭龍類。第二次滅絕事件是三疊紀-侏儸紀滅絕事件,發生於約2億年前,大部分的早期主龍類滅亡,包含:堅蜥類、鳥鱷科、植龍目、與勞氏鱷類。這群動物滅亡後,鱷形超目、恐龍、哺乳類、翼龍目、與烏龜開始佔據陸地的生態系統[20]。

數種三疊紀恐龍的化石:艾雷拉龍(大)、始盜龍(小)、以及板龍的頭顱骨(右)
原始恐龍的第一次適應演化發生於三疊紀的卡尼階與諾利階之間,極可能因為當時有許多優勢動物滅亡。在傳統的觀念裡,在三疊紀中期,恐龍與當時其他的陸地動物展開長時間的競爭,並取得優勢,但這是不可能的。根據化石紀錄,恐龍沒那麼早出現,且早期恐龍並非穩定的增加、多樣化,而是後來才急驟的大量演化;如果牠們與其他動物有過長時間競爭,應該會在化石紀錄中反映出來。恐龍在卡尼階時的數量非常稀少,僅佔陸地動物群的1~2%。在諾利階,數個動物群滅亡後,恐龍則佔了陸地動物群的50~90%。恐龍的直立步態過去被視為成功演化的關鍵,但現在已知有數個同時代的動物群也有直立步態,例如:堅蜥類、鳥鱷科、勞氏鱷類、以及數種鱷形超目動物。三疊紀後期是個生物劇變的時代,有許多植物、海生動物興起衰落,三疊紀後期的氣候也有巨大改變[20]。鑲嵌踝類主龍目前僅剩下鱷魚,但在三疊紀晚期,鑲嵌踝類主龍是群非常多樣化的演化支,包含:鳥鱷科、堅蜥目、植龍目、勞氏鱷目。在這群優勢物種存在下,恐龍存活到侏羅紀,並非生理、競爭上佔有優勢,而是面對滅絕事件的運氣問題[39]。
科学分类
恐龍與現代鳥類、鱷魚都屬於主龍類。大部分的爬行動物與鳥類,都屬於雙孔動物;哺乳類與似哺乳爬行動物擁有一個顳顬孔,合稱合弓動物;烏龜與早期的副爬行動物,沒有顳顬孔,則屬於無孔動物。恐龍與其他主龍類擁有許多生理構造上的特徵,例如:牙齒位於齒槽中,而非直接從顎部骨頭延伸。在主龍類中,恐龍的最顯著特徵是牠們的步態。恐龍的後肢直立於身體下方,而蜥蜴與鱷魚的四肢則是往兩側攤開。
恐龍指的是特定陸地爬行動物,能採取直立步態,包括蜥臀目與鳥臀目,並不包括翼龍類、魚龍類、蛇頸龍類、滄龍類[28]。恐龍常被認為是總目,或是未定位的演化支。蜥臀目和鳥臀目以其骨盆结构來区分。
蜥臀目(Saurischia)意為「蜥蜴的髖關節」,骨盆型態比較接近早期的祖先,恥骨朝前[25]。數個演化支的恥骨個別演化成朝向後方,例如:艾雷拉龍下目[40]、鐮刀龍超科[41]、馳龍科[42]、以及其後代鳥類[30]。蜥臀目主要包括兩個亞目:二足、大部分是肉食性的獸腳亞目;長頸部、四足、草食性的蜥腳形亞目。某些蜥臀目大約在侏儸紀時期演變成為鳥類的祖先。
鳥臀目(Ornithischia)意為「鳥類的髖關節」,擁有類似鳥類的骨盆結構,恥骨朝向後方,但這是趨同演化的結果。與鳥類不同的是,鳥臀目的恥骨有個朝前的突出部分。鳥臀目包含多樣性的草食性動物。
根據定義,蜥臀目的範圍是:與鳥類具有最近共同祖先,而與鳥臀目具有較遠共同祖先的所有恐龍;鳥臀目的範圍則是:與三角龍具有最近共同祖先,而與蜥臀目具有較遠共同祖先的所有恐龍。
.mw-parser-output .gallery-mod{background:transparent;margin-top:0.5em}.mw-parser-output .gallery-mod-collapsible{width:100%}.mw-parser-output .gallery-mod-center{margin:0 auto}.mw-parser-output .gallery-mod-title{text-align:center;font-weight:bold}.mw-parser-output .gallery-mod-box{float:left;border-collapse:collapse;margin:3px}.mw-parser-output .gallery-mod-box .thumb{border:1px solid #ccc;background-color:#F8F8F8;padding:0;text-align:center}.mw-parser-output tr.gallery-mod-text{vertical-align:top}.mw-parser-output tr.gallery-mod-text .core{display:block;font-size:small;padding:0}.mw-parser-output .gallery-mod-text .caption{line-height:1.25em;padding:6px 6px 1px 6px;margin:0;border:none;border-width:0;text-align:left}.mw-parser-output .gallery-mod-footer{text-align:right;font-size:80%;line-height:1em}
|




| 蜥臀目 | 獸腳亞目 | 坚尾龙类 | 棘龙超科:斑龙科、棘龙科 肉食龙下目:单脊龙、异特龙超科 虚骨龙类 右图从左到右为:棘龙、鲨齿龙、南方巨兽龙、暴龙 | 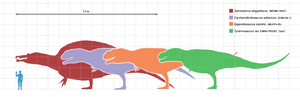 | |
| 蜥腳形亞目 | 原蜥脚下目 | 里奧哈龍科(優脛龍、里奧哈龍)、板龍科(板龍、鞍龍、黑水龍)、大椎龍科(科羅拉多斯龍、祿豐龍、大椎龍、雲南龍、金山龍) |  | ||
| 蜥脚下目 | 近蜥龍科、黑丘龍科、貝里肯龍科、火山齒龍科、鯨龍科、馬門溪龍科、圖里亞龍類 | ||||
| 新蜥腳類 | 梁龙超科:雷巴齊斯龍科、叉龍科、梁龍科(迷惑龍、梁龍) | ||||
大鼻龙类 右图是數種大鼻龍類恐龍,由左到右依序為:圓頂龍、腕龍、長頸巨龍、盤足龍。 | |||||
| 鳥臀目 | 装甲类 | 腿龍科、劍龍下目、甲龍下目 | |||
| 角足亚目 | 鳥腳下目 | 稜齒龍科、禽龍科 鴨嘴龍超科:慈母龙、鸭嘴龙 右图:左後:彎龍、左:禽龍、中後:山東龍、中前:橡樹龍、右:冠龍、右後(小):畸齒龍、右後(大):腱龍 | 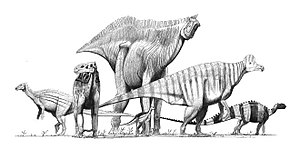 | ||
| 頭飾龍類 | 厚頭龍下目、角龍下目 | ||||
演化與地理分佈
在三疊紀之後,恐龍的演化隨着植被與大陸位置的改變而變動。在晚三疊紀到早侏儸紀期間,各主要大陸連接成一個超級大陸,盤古大陸。這段時間的恐龍動物群主要由肉食性的腔骨龍超科、草食性的原蜥腳下目所構成[43]。裸子植物(尤其是松柏目),在晚三疊紀開始輻射演化,是當時的可能食物來源之一。原蜥腳類恐龍的頜部只能撕咬下樹葉,但不能做出複雜的嘴部動作,因此必須要藉由其他的方式協助磨碎食物[44]。
恐龍動物群維持了一段同質性,直到侏儸紀中到晚期,此時的掠食性恐龍開始多樣化,例如角鼻龍下目、棘龍超科、肉食龍下目,草食性恐龍則有劍龍下目、體型巨大的蜥腳下目。這段時間的著名地層有:北美洲的莫里遜組、坦尚尼亞的敦達古魯地層、中國的大山鋪組。此時的中國地區,也呈現出多樣性的恐龍,例如中華盜龍科、頸部極長的馬門溪龍[43]。甲龍下目與鳥腳下目開始普遍,而原蜥腳類恐龍則已滅絕。松柏目與蕨類成為最普遍的植物。蜥腳類恐龍與早期的原蜥腳類恐龍類似,嘴部無法做出複雜的動作。鳥腳類恐龍則發展出不同的處理植物方式,例如可將食物置在嘴中的兩側頰部,以及類似咀嚼的頜部動作[44]。這段期間的另一個重要演化事件是鳥類的演化出現,牠們是從虛骨龍類的手盜龍類演化而來[30]。

18種原始角龍類的體型比較
左排由上到下為:隱龍、遼寧角龍、古角龍、祖尼角龍、鬥吻角龍、纖角龍。
中排由上到下為:朝陽龍、蒙古鸚鵡嘴龍、閻王角龍、雅角龍、安德薩角龍、蒙大拿角龍。
右排由上到下為:黎明角龍、紅山龍、西伯利亞鸚鵡嘴龍、原角龍、弱角龍、傾角龍
到了早白堊紀,盤古大陸持續的分裂成各陸塊,恐龍變得更多樣化。甲龍類、禽龍類、腕龍科散佈於歐洲、北美洲、以及北非。稍後,大型獸腳亞目恐龍,例如棘龍科、鯊齒龍科,以及蜥腳下目的雷巴齊斯龍科與泰坦巨龍類,佔據者非洲與南美洲。在亞洲,手盜龍類的馳龍科、傷齒龍科、偷蛋龍下目成為常見的獸腳類恐龍,甲龍科與早期角龍下目(例如鸚鵡嘴龍)則是該地常見的草食性動物。在同一時期,澳洲則生存者原始的甲龍類、棱齒龍類、禽龍類[43]。劍龍類則在早白堊紀到晚白堊紀之間滅絕。早白堊紀發生了一件重大變化,開花植物出現在北美洲,並在晚白堊紀擴散分布。數群草食性恐龍發展出不同的方式以協助進食。角龍類發展出切割用的喙嘴,後方有眾多咀嚼用齒系(Tooth batteries);禽龍類也獨自演化出咀嚼用齒系,並在鴨嘴龍科發展到極致[44]。少部份蜥腳類恐龍也演化出齒系,雷巴齊斯龍科的尼日龍是最明確的證據[45]。
晚白堊紀的恐龍通常分為三個動物群。在北半球的北美洲與亞洲,獸腳類恐龍主要有大型的暴龍科、以及眾多的小型手盜龍類恐龍,優勢草食性恐龍則由鴨嘴龍科、角龍下目、甲龍科、厚頭龍下目所構成。南半球的岡瓦那大陸持續分裂中,阿貝力龍科是優勢獸腳類恐龍,泰坦巨龍類則是優勢草食性恐龍。在歐洲,凹齒龍科、結節龍科、泰坦巨龍類則是常見的恐龍[43]。開花植物更為輻射演化[44],第一群禾本科(草)植物出現在白堊紀末期[46]。鴨嘴龍科與甲龍下目在北美洲、亞洲發展至極度多樣化。獸腳亞目也演化出草食性或雜食性的物種,鐮刀龍超科與似鳥龍下目更為常見[44]。
白堊紀-第三紀滅絕事件發生於白堊紀末期,接近6500萬年前,造成所有恐龍的滅絕,以及鳥類的崛起。但仍有其他雙孔動物在滅絕事件中存活下來,例如:鱷魚、蜥蜴、蛇、喙頭蜥、以及離龍類[47]。
白堊紀晚期的多樣性降低?
在傳統的觀念裡,在白堊紀晚期,恐龍的生物多樣性達到最高,發展出最多的物種。在2008年,Graeme T. Lloyd等人提出這是化石紀錄取樣偏差所造成的假象,由於白堊紀晚期的地層較容易被發現,因此最容易挖掘出這個時期的化石,並不代表白堊紀晚期的恐龍最具多樣性。Lloyd等人更提出,這個時期只有兩個恐龍演化支持續演化輻射出高度多樣性,例如:鴨嘴龍類、角龍類。在白堊紀早期以前,陸地的優勢植物是裸子植物,例如針葉樹;在白堊紀中晚期,開花植物開始成為優勢植物之一,但其地理範圍有限。根據恐龍的糞化石顯示,白堊紀晚期的大部分草食性恐龍仍以裸子植物為食,只有少部分以開花植物為食。同時,草食性昆蟲、哺乳動物的物種數量迅速增加,顯示牠們已經適應以這種新型態植物為食。除此之外,當時的蜥蜴、蛇、鱷形類、以及鳥類都出現多樣性增加的跡象。Lloyd等人假設,相對於這些動物的多樣性增加,恐龍的多樣性則降低,這可能是導因於緩慢變遷的植物生態系統[48]。
古生物學
恐龍的相關知識來自於多樣化的非化石與化石紀錄,例如:化石、糞化石、足跡化石、胃石、羽毛、皮膚痕跡、內臟、與軟組織[49][50]。物理學(尤其是生物力學)、化學、生物學、地球科學等相關研究也增長了恐龍的相關知識。恐龍研究的最重要兩個課題是恐龍的體型與行為。
體型

来自四個恐龍亞目的大型代表物種體型相比。前面從左到右分別為:巨型山東龍(紅)、埃及棘龙(綠)、蹄足剑龙(橘)、恐怖三角龙(蓝)以及后方的烏因庫爾阿根廷龍(紫),圖片最左邊淺藍色的則代表現代人類的平均體型。
尽管目前证据不全,恐龙整体而言的体型很大,當然這是因為當時地球環境保持長時間的穩定與濕熱的緣故,而非現在的人類所處的冰河期。以恐龙作為标准来看,蜥脚下目是其中的巨无霸。在漫长的恐龙时代,即使是体型最小的蜥脚類恐龍也要比牠們栖息地內的其他动物要大,而最大的蜥脚类则比任何出現在地表的动物都要大出几个等级。巨型史前哺乳动物,如巨犀和猛犸,在巨型蜥脚類动物前不值一提,仅有少数的现代海洋生物的體型接近或超过大型蜥腳類恐龍:最为人熟知的是蓝鲸,體重可以达到173公噸,身長超過30米长[51]。
蜥腳類恐龍的巨大體型,可能具有幾項優勢,例如防禦掠食動物、減低能量的使用、長壽,其中最重要的因素應為消化食物。與小型動物相比,大型動物的消化作用更具效率,因為食物在牠們的消化系統中待更多的時間。基於此原因,大型動物可用較低營養價值的食物維持生存。蜥腳類恐龍的化石,大部分發現於過去為乾旱或季節性乾旱的地層中,牠們可以大量、低營養價值的植物為食,以在這些環境中維持生存[52]。
然而,大部分的恐龙在時間尺度上還是要比大型蜥腳類恐龍還小得多。现有的证据表明,恐龙的平均大小在三叠纪、早侏罗纪、晚侏羅紀和白垩纪都不断变化[37]。大部分獸腳類恐龍的體重在100到1,000公斤之間,而全新世的掠食性肉食動物則多半在10到100公斤之間[53]。恐龍的體重估計值,大多介於1到10公噸之間[54]。倫敦國立自然歷史博物館的一個研究指出恐龍的體重平均值約在100公斤左右,而新生代的哺乳類體重平均值多在2到5公斤之間[55]。
最大和最小的恐龙
只有很小比例的动物最后会成为化石保存下来,况且绝大部分的化石都仍埋藏于地底。只有极少数的已被发现的样本保留有完整的骨骼,而其中保存有皮肤和其他软组织痕跡的则更为稀有。藉由比較不同骨頭的大小與型態,来重建一副完整的骨架,是一個複雜的過程,而重塑肌肉和器官则更為困難。綜合以上幾點理由,科学家可能永远无法确定最大和最小的恐龙。

五種最大型的恐龍體型比較,從左至右分別為烏因庫爾阿根廷龍(紅)、中加馬門溪龍(綠)、薇薇安超龍(橘)、哈氏梁龍(紫,即地震龍)以及杜氏富塔隆柯龍(藍)

四種最小型的非鳥獸腳恐龍體型比較,從左至右分別為分類存疑的「湯氏古老翼鳥龍」(橘)和「微小似鳥龍」(紫)、遙遠小馳龍(紅)以及胡氏耀龍(綠)
从现有的状况较好的骨架来看,最高和最重的恐龙是長頸巨龍(Giraffatitan brancai,原名布氏腕龍)。長頸巨龍的化石在1907年至1912年间发现于坦桑尼亚。从多具大小相近的个体所組合而成的骨架模型,现正展示于柏林洪堡博物馆[56],该模型高12米,长22.5米,预期活体的重量在30到60公噸之间。最长的恐龙是27米长的梁龙,是在1907年发现于美国怀俄明州,現正展示于賓州匹兹堡的卡内基自然历史博物馆。
但是对於这些巨大的恐龙的所有知識,都是根據少數碎乱的化石而来。化石记录中的最巨型草食性恐龍標本,大部分是在1970年代后被发现的,包括重大的阿根廷龙,体重估計在80到100公噸之间;梁龍的一個種(Diplodocus hallorum,原名地震龍)身長33.5公尺[52],而超龙身長33公尺长[57];最高的恐龍是18米高的波塞东龙,頭部可以達到6层楼的窗口。而易碎雙腔龍(Amphicoelias fragillimus)可能是最长的恐龍,但是对雙腔龍的全部知識都来自于1878年發現的一個脊椎神經棘化石,该化石现已遗失。从当时对该化石的图释推断,它有可能达到58米长和120公噸重[52]。雙腔龍的重量仅次于罕为人知的巨体龙,巨體龍的体重可能达到175到220公噸。
目前已知最長的肉食性恐龍是棘龍,身長為16到18公尺,重量為8,150公斤[58]其他的巨型食肉性恐龍有南方巨獸龍、马普龙、暴龙、和鲨齿龙。
不计现代鸟类(如:吸蜜蜂鸟)的话,最小的恐龙体型相当于鴿子[59]。兽脚亚目的近鳥龍和耀龍体长都在35公分以下[60][59]。目前已知最小型的成年恐龍標本屬於近鳥龍,體重估計為110公克左右[60]。而已知最小型的草食性恐龍則是微角龍與皖南龍,身長約60公分[61][62]。
行為

在1978年發現的慈母龍,具有築巢的行為
對於恐龍行為的假設,多是根據化石姿態、棲息地、電腦模擬的生物力學、以及與類似生態位的現代動物比較而來。這些行為假設多存在著爭議。目前科學家們普遍同意鱷魚與鳥類所共同擁有的行為,有可能也存在於牠們的近親恐龍身上。
在1878年,比利時貝尼沙特發現了31個禽龍化石,可能是洪水所造成的集體死亡,這是恐龍群居行為的第一個證據[63]。之後在其他的地層也發現集體死亡的現象,再加上群集的足跡化石,顯示許多恐龍物種具有群居的行為。目前已發現由數百個、甚至數千個個體所留下的鴨嘴龍科足跡化石,顯示鴨嘴龍科可能會大量群體移動,如同今日的美洲野牛與非洲跳羚。在英格蘭牛津發現的蜥腳類足跡化石,顯示不同的蜥腳類恐龍會共同集體移動[64],但是沒有證據可辨認出此足跡化石由哪些恐龍所留下[65]。恐龍群體行動可能有許多原因,例如防禦掠食動物、集體遷徙,或者保護幼年個體。有證據顯示部分恐龍的未成年個體會群體行動,例如:獸腳亞目、蜥腳下目、甲龍下目、鳥腳下目、角龍下目。舉例而言,中國內蒙古曾發現20多個群聚的中國似鳥龍化石,年齡為1到7歲,科學家認為這些中國似鳥龍是集體行動時,陷在泥濘中死亡[66]。某些肉食性的獸腳亞目恐龍,也曾被認為會以小群體方式行動,以獵食體型更大的獵物[67][68]。但是,恐龍的現代近親鱷魚、其他爬行動物、與鳥類,多半沒有群體獵食的行為,栗翅鷹是個少數例外。埋葬學證據顯示恐爪龍與異特龍的集體化石,可能是爭奪食物時打鬥造成的死亡,如同許多現代的掠食性爬行動物,例如科摩多龍[69]。

偷蛋龍科葬火龍的蛋巢,位於紐約美國自然歷史博物館
在1978年,古生物學家傑克·霍納(Jack Horner)於蒙大拿州發現慈母龍的化石與蛋巢遺跡,顯示鳥腳下目具有長時間的親代養育行為[70]。在1997年,在巴塔哥尼亞白堊紀地層發現的泰坦巨龍類化石,顯示這些大型恐龍也具有築巢的行為[71]。在1993年,在蒙古發現的偷蛋龍科葬火龍化石,被發現採取類似雞的孵蛋姿勢,牠們被推測身體可能覆蓋者羽毛,以保持蛋的溫度[72]。一些證據顯示某些恐龍可能具有親代養育行為。例如,由一個成年體、34個幼年體構成的鸚鵡嘴龍群體化石,其大量的幼年體比例,可能就是親代養育的證據[73]。此外,一個大椎龍的胎胚化石,被發現沒有牙齒,顯示牠們在孵化過後,仍有一段時間需要親代餵食[74]。在蘇格蘭西北方斯開島發現的足跡化石,也指出鳥腳下目有親代養育行為[75]。許多的恐龍主要演化支都已發現蛋巢與蛋化石,恐龍可能與其幼年體之間有某些溝通行為,類似現代鳥類與鱷魚。

兩隻打鬥中的尖角龍,牠們生存於晚白堊紀的北美洲
某些恐龍具有頭冠或是頭盾,例如頭飾龍類、獸腳亞目、以及賴氏龍亞科,牠們的頭冠、頭盾過於脆弱,無法用來自我防衛,而可能具有性展示功能,或是使用在物種內打鬥行為;但目前對於恐龍的求偶與領域性行為所知有限。一些瘦腳亞目化石的頭部具有傷口,可能是同類互相攻擊的後果[76]。恐龍的溝通方式仍充滿謎題,是個正在研究中的領域。近年的研究顯示,賴氏龍亞科的中空頭冠可能具有揚聲器作用,可用來與同類之間做聲音的溝通、辨認[77][78]。
在1971年,在蒙古戈壁沙漠發現了一個極為著名的化石標本,一隻伶盜龍正在攻擊一隻原角龍[79],這是恐龍會獵食其他恐龍的直接證據[80]。另外,一個埃德蒙頓龍的尾巴有個可能的瘉合痕跡,可能是由暴龍科恐龍所留下[80]。在2003年,在馬達加斯加所發現的瑪君龍化石,其骨頭上有同類的齒痕,證實某些獸腳類恐龍具有同類相食行為[81]。
根據目前的化石證據,某些草食性恐龍可能是穴居動物,例如稜齒龍類的掘奔龍[82];而某些獸腳類恐龍則是樹棲動物,例如原始的馳龍科小盜龍[83]、與充滿謎題的擅攀鳥龍科[84]。但大部分恐龍則是依靠陸地為生存。生物力學可以用來研究恐龍的移動方式,進而研究牠們的相關行為模式。藉由計算恐龍的肌肉力量傳遞模式,以及重心位置,可推算出恐龍的移動速度[85]。此外,生物力學還可計算出梁龍科的鞭狀尾巴揮動時是否能產生音爆[86]、以及蜥腳類恐龍能否在水中浮動[87]。
生理特徵

暴龍的雙眼朝前,具有立體視覺。位於巴黎探索皇宮

Eubrontes,一個發現於猶他州西南部蒙納夫組(下侏儸紀)的足跡化石。
自從60年代以來,恐龍的體溫調節成為科學家們爭論的主題之一。最初,科學家們對於恐龍的體溫調節能力的看法相當不一致。近年來,恐龍恆溫動物說逐漸成主流理論,牠們被視為活躍的動物,至少具有相當穩定的體溫。目前的爭論多在於牠們的體溫調節機制,以及牠們與鳥類、哺乳類的代謝率相近程度。
在恐龍研究的早期,古生物學家認為恐龍是類似蜥蜴的冷血動物。他們認為恐龍是群相當緩慢、慵懶的動物,需要外部熱量以調節牠們的體溫,接近現代爬行動物。恐龍冷血動物說成為主流看法,直到1968年,羅伯特·巴克公佈了一份革命性的研究,提出恐龍溫血動物說。
目前的證據多指出恐龍可生存在較寒冷的氣候環境,且至少有某些恐龍藉由內部身體機制來調節體溫,例如巨大體型造成的巨溫性。恐龍恆溫動物說的證據包含:在南極洲與澳洲發現恐龍化石,兩地區在當時位於極區內,有長達六個月的永夜、某些恐龍具有隔絕熱量用的羽毛、以及骨頭內具有恆溫動物特有的心血管結構。從骨骼結構顯示,獸腳類與其他恐龍是活躍的動物,若牠們具有恆溫動物的心血管系統,將符合這樣的生活方式;而蜥腳類恐龍則擁有較少的恆溫動物體徵。最有可能的狀況是,某些恐龍是恆溫動物,而其他則不是。恐龍恆溫/變溫動物說的爭論與新證據仍在持續進行中[88]。
因為有許多特徵可用來判斷是否為恆血動物,使得恐龍恆溫/變溫動物說的爭論難以定論。大部分的研究人員將牠們與鳥類、哺乳類相比。小型的鳥類、哺乳類的身體覆蓋者隔絕物,例如:脂肪、皮毛、羽毛,可減緩身體熱量的流失。大型鳥類、哺乳類的身體表面積/體積比例較低。根據霍爾丹法則(Haldane's principle),與身體體積相比,大型動物反而擁有較小比例的表面積,牠們釋放的熱量溫度較高,而吸收的熱量溫度較低。在某種程度體型以上,身體所散發的熱量,低於身體所製造的熱量,這將使得動物採用其他方式來散發熱量。以大象為例,牠們皮膚缺乏毛髮,但有大型耳朵,可增加身體表面積,或者將身體浸泡在水或泥濘中來降低體溫。
大型恐龍被認為具有相同的狀況,牠們的巨大體型使熱量散失較慢,體溫高於所處環境,造成巨溫性。但這仍無法解釋眾多的小型恐龍、或幼年個體的體溫調節機能。
在2000年,一個對於奇異龍標本的電腦斷層掃描,發現了可能是四腔室心臟的結構,類似現代鳥類與哺乳類[89]。這個研究在科學界中引起爭議,有其他生理結構[90]、或其他物體等反對意見[91]。恐龍的近親,現代鱷魚與鳥類都擁有四腔室心臟(鱷魚的心臟較不一樣),所以恐龍可能也有類似的心臟結構;但心臟結構與代謝率沒有必然的關係[92]。
軟組織與DNA

棒爪龍的化石具有軟組織痕跡,位於義大利米蘭米蘭自然歷史博物館
在1998年,義大利的一處石灰岩地層出土了一個虛骨龍類恐龍化石,名為棒爪龍,這個幼年體化石保存了極為良好的軟組織痕跡,包含部份氣管、小腸、結腸、肝臟及肌肉[49]。
在2005年五月份的《自然》雜誌中,北卡羅萊納州立大學的瑪莉·海格比·史威茲(Mary Higby Schweitzer)與其同事,宣佈在一個暴龍腿部骨頭的骨髓中發現了軟組織。這個化石發現於蒙大拿州海爾河組,年代為6,800萬年前[50]。
經過數週的去礦化處理後,史威茲與其團隊等人鑒定出分叉的血管,以及纖維狀的骨頭組織。此外,骨頭組織中還具有類似血球細胞的微小組織。這個骨頭的結構類似鴕鳥的血球細胞與血管。關於這些組織的真實身分,研究人員目前還沒有確定地做出定論。相關的研究仍在進行中[50]。
在2008年7月份的《公共科學圖書館·綜合》(PLoS One)雜誌中,華盛頓大學的凱伊·托馬斯(Thomas Kaye)等人對上述的暴龍軟組織提出質疑。他們認為這些骨頭中的軟組織僅是細菌形成的生物薄膜,這些細菌生存在原本血管與細胞的空間[93]。托馬斯等人認為這些結構被誤認為是血球細胞,是因為帶有鐵的微球粒存在。他們在許多不同時期的其他化石,發現類似的微球粒,例如菊石。在菊石的案例中,他們發現含有鐵的空間,與血液沒有直接的關連[94]。
目前已有兩個團隊,宣稱已在恐龍化石中成功擷取出DNA,但根據更進一步的檢驗與同行評審,兩者的發現都無法受到肯定[95]。根據一份2002年的研究,藉由比較現代爬行動物與鳥類的視色素中的胺,可用來推算、重建出原始主龍類與恐龍的胺基酸序列,甚至是基因序列[96]。此外,已在恐龍化石中發現數種可能是蛋白質的物質[97],包含血紅素[98]。
即使恐龍的DNA可以重建,以目前的科學技術,想要重建出一隻恐龍並培養長大,是極度困難的。原因是目前沒有與恐龍極相近的物種可供受精,也沒有適合的環境可使胚胎發育。
與鳥類的關係
恐龍可能是鳥類祖先的理論,是在1868年由湯瑪斯·亨利·赫胥黎首次提出[99]。在20世紀早期,格哈德·海爾曼(Gerhard Heilmann)出版了一本書,《鳥類起源》(The Origin of Birds)。海爾曼根據恐龍缺乏叉骨(接合的鎖骨),而認為鳥類與恐龍間沒有關係,他進而假設鳥類演化自鱷形超目或槽齒目的祖先,而非恐龍[100]。後來發現的恐龍化石則發現了鎖骨或叉骨[30],例如1924年發現的偷蛋龍,但在當時被誤認為是間鎖骨[101]。在1970年代,約翰·奧斯特倫姆(John Ostrom)重新提出鳥類演化自恐龍的理論[102]。隨着這幾十年來親緣分支分類法研究的增加,以及更多小型獸腳類恐龍與早期鳥類的發現[21],這個理論得到更多的支持[103]。其中最重要的是發現於義縣組的有羽毛恐龍化石,已發現過多種不同型態的羽毛[30]。鳥類與獸腳類恐龍具有上百個共同的生理特徵,牠們目前被普遍認為是近親[104]。鳥類與手盜龍類的關係更為親近[30]。少數科學家則提出不同的演化途徑,例如艾倫·費都加(Alan Feduccia)與賴瑞·馬丁(Larry Martin),他們提出類似海爾曼的鳥類演化自原始主龍類假設[105],或是鳥類起源於手盜龍類,但手盜龍類並非恐龍,而是與恐龍的趨同演化結果[5]。
有羽毛恐龍

著名的印石板始祖鳥標本,位於德國。
始祖鳥發現於1861年的德國南部,是第一個所謂的「有羽毛恐龍」。這個標本出土於索倫霍芬石灰岩礦床,這個地層因為挖出了許多狀態極良好的化石而著名。始祖鳥是個過渡化石,明顯具有現代爬蟲類與鳥類的中間特徵。而就在兩年前,查爾斯·達爾文發表了《物種起源》一書,始祖鳥的發現使得演化論與創造論的爭論更為激烈。這種早期鳥類非常像恐龍,至少有一個標本,因為周圍沒有明確的羽毛痕跡,而被誤認為是美頜龍[106]。
90年代以來,發現了更多的有羽毛恐龍,為恐龍與鳥類的關係提供了更多的證據。大部分的新標本被發現於中國遼寧省的義縣組,該地在白堊紀的時候,屬於一個小型大陸。雖然有羽毛恐龍僅發現於少數地區,但有可能當時全球各地都出現了有羽毛恐龍。各地的化石紀錄很少羽毛痕跡與有羽毛恐龍,可能是因為在化石化的過程中,羽毛與皮膚等脆弱部份,很難被保留下來。原始的羽毛細而缺乏羽枝,已出現在原始的虛骨龍類恐龍,例如美頜龍科的中華龍鳥,與暴龍超科的帝龍[107],而有羽枝的羽毛出現在虛骨龍類的手盜龍類,例如:偷蛋龍下目、傷齒龍科、馳龍科、以及鳥類[30][108]。但是,有羽毛恐龍本身也有一些爭議,其中最重要的是艾倫·費都加(Alan Feduccia)與蒂綱·索拉(Theagarten Lingham-Soliar)所提出的,他們指出這些看起來是原始羽毛的物體,其實是膠原纖維腐化而成的,這些膠原纖維在身前位於恐龍外表覆蓋物之下[109][110][111]。他們並提出手盜龍類都不是恐龍,而是與恐龍平行演化的結果[5][110]。但他們的大部分意見不被其他研究人員所接受[112]。
骨骸
因為羽毛被認為與鳥類有關聯,有羽毛恐龍被視為是恐龍與鳥類之間的失落環節。除此之外,恐龍與鳥類之間還有許多骨骼上的共同特徵,例如:頸部、恥骨、腕骨、手臂、肩帶、叉骨、以及龍骨。藉由親緣分支分類法研究,兩者之間的共同骨骼特徵,加強了恐龍與鳥類的關係與連結。

氣腔龍的各部位骨頭,呈現許多空洞空間
軟組織
根據俄亥俄大學的古生物學家派崔克·歐康納的研究,大型肉食性恐龍具有複雜的空腔,類似現代鳥類的氣囊。這些腳踝類似鳥類的二足獸腳類恐龍,其肺臟可將空氣送往骨頭內的空腔,如同現代鳥類。氣囊過去被認為是鳥類的專有特徵,但目前已發現某些恐龍具有類似的氣囊系統[113]。在2008年7月份的《公共科學圖書館·綜合》(PLoS One)雜誌中,科學家宣稱發現一種新恐龍,氣腔龍,具有類似鳥類的呼吸系統。電腦斷層掃描顯示氣腔龍骨頭的內部具有中空空間[114][115]。
恐龍與鳥類有接近親緣關係的另一個證據則是沙囊石頭。這些動物會吞食石頭以協助消化,並磨碎胃中的食物與堅硬纖維。發現於化石中的沙囊石頭,被稱為胃石[116]。
繁殖
一個近幾年發現的暴龍化石,其腿部骨頭被發現有骨髓骨。這被視為恐龍與鳥類演化自共同祖先的證據,也是首個可鑑定出性別的恐龍化石。雌性鳥類在產卵時,四肢骨頭的外部骨質與骨髓之間會產生一種特殊骨層,稱為骨髓骨(Medullary bone)。骨髓骨富含鈣,只存在於鳥類體內,可在產卵期用來製造蛋殼[117]。目前已在獸腳亞目的暴龍與異特龍、以及鳥腳下目的腱龍化石上發現骨髓骨。因為這三者與鳥類的所屬支系,在相當早的時候就已分開演化,因此可推論恐龍普遍會製造骨髓組織。由於曾在亞成年體化石發現過骨髓骨,這顯示恐龍達到性成熟的時間很快[118]。
行為證據
一個近年發現的傷齒龍科化石,顯示牠們睡覺時,頭部塞在手臂內側,類似某些現代鳥類[119]。這種睡姿可使頭部保持溫暖,是現代鳥類的特徵之一。
灭绝理论
非鳥類恐龙消失于6千5百万年前的白垩纪-第三纪灭绝事件,同時間還有許多生物也跟著滅亡,包含:菊石、滄龍科、蛇頸龍目、翼龍目、草食性的烏龜與鱷魚、大部分鳥類、以及原始哺乳類[14]。自從1970年代以來,有許多研究試圖探索這次滅絕事件的原因,並形成數個理論。通常的解释是一個撞擊事件造成了灭绝,有些科學家提出其他理由,或者是幾個綜合事件導致這次滅亡。
在恐龍時代末期,極區並沒有冰帽,海平面比現在高出100到250公尺。地球各地的氣溫差異不大,極區與赤道地區的氣候相差不到25°;整體而言較今日暖和,極區的氣溫高於現在的氣溫約50°[120][121]。
恐龍時代的大氣成分也與今日有極大差別。二氧化碳是今日的12倍,氧氣則佔了大氣的32%到35%,而今日只佔了21%。但是在晚白堊紀,地球的環境有顯著的變動。火山活動增加,造成氣候變冷,二氧化碳比例下降,氧氣比例開始變動。有些科學家認為氣候變化、低氧氣比例,可能是滅亡的直接原因。如果恐龍的呼吸系統類似現代鳥類,牠們的呼吸效能將會有困難,需要更多氧氣才能維持巨大體型的需要[14]。

位於猶加敦半島的希克蘇魯伯隕石坑,這次撞擊事件可能導致恐龍的滅亡。
撞击事件
在1970年代後期,加州大學柏克萊分校的路易斯·阿尔瓦雷茨提出了彗星撞擊理論,造成了白堊紀末期的大規模滅絕事件。阿爾瓦雷茨提出世界各地的地層,銥含量在6,500萬年前有大量的增加,可視為撞擊的直接證據[122]。有許多證據顯示當時有顆直徑5到15公里的隕石或彗星,撞擊現在的猶加敦半島地區,形成寬170公里的希克蘇魯伯隕石坑,引發此次滅絕事件[123][124]。但科學家不確定恐龍是在撞擊事件中滅亡,還是在此之前就已經衰退。另有科學家提出這次撞擊,使大氣層佈滿灰塵,或者是使地球氣溫上升。
化石紀錄不能看出恐龍滅絕的速度,但許多不同的測試研究發現牠們的滅亡相當迅速。這些結果使科學界更加相信彗星撞擊理論。撞擊事件短期內會造成高溫,而揚塵會遮闢天空,造成全球性的長時間氣候冷卻。大部分植物因無法光合作用而消失,草食性恐龍也因沒有食物而死亡,而肉食性恐龍也因沒有食物來源,也漸漸滅亡,僅有小型動物和少量植物倖存下來。
在2007年9月,美國科羅拉多州美國西南研究院的研究人員威廉·波特克(William Bottke)與數位捷克籍的科學家,利用電腦計算出希克蘇魯伯撞擊事件的可能肇事者。根據他們的模擬計算結果,該隕石坑有90%的可能性是由一個名為巴普提斯蒂娜的小行星所造成的,該小行星直徑接近160公里,運行於火星與木星之間;在1億6000萬年前,巴普提斯蒂娜小行星被一顆直徑約55公里的未命名小行星撞擊後粉碎,主要的碎片形成巴普提斯蒂娜族小行星帶,而某些碎片則闖入地球的公轉軌道,其中一顆直徑10公里的碎片,在6,500萬年前撞擊了墨西哥猶加敦半島,形成希克蘇魯伯隕石坑[125]。
威廉·波特克的研究結果與阿爾瓦雷茨的理論相似,另外也採納了柏克萊大學的物理學家查理·穆勒所提出的太陽伴星理論。查理·穆勒提出太陽有顆名為復仇女神(Nemesis)的伴星,復仇女神每隔2,600萬年會經過奧爾特雲彗星帶,干擾了彗星的軌道,使眾多彗星進入內太陽系[126]。這些彗星的少部份會進入地球的公轉軌道,與地球發生撞擊,進而造成大規模的滅絕事件[126]。
火山爆發
在2000年以前,就有學者主張德干暗色岩(Deccan Traps,一種洪流玄武岩地形)是這次滅絕事件的主因,但因為這次火山爆發始於6,800萬年前,並持續超過200萬年,所以當時認為德干暗色岩火山爆發所造成的滅絕事件應該是逐步、緩慢的。但現在有證據顯示,有2/3的德干暗色岩地形,是在一萬年內形成的,發生於6,550萬年前。顯示這次火山爆發造成了快速的滅絕事件,可能在數千年內,但還是遠比撞擊事件長[127][128]。
德干暗色岩火山爆發藉由數種方式造成了滅絕事件,灰塵與二氧化硫與二氧化碳等溫室氣體大量噴出,灰塵使地表的日照量下降,植物的光合作用減少;當灰塵慢慢降落至地表時,火山爆發噴出的二氧化碳氣體,可能增加了溫室效應。許多植物因此而死亡,草食性恐龍因沒有食物而滅亡,而肉食性恐龍也相繼滅絕[128]。在恐龍大規模滅絕前,火山爆發噴出的氣體,似乎與全球暖化有關。有證據指出在希克蘇魯伯撞擊事件之前的50萬年內,大氣溫度曾上升8 ℃[127][128]。
在德干暗色岩火山爆發仍被認為是緩慢滅絕事件時,路易斯·阿爾瓦雷茨便已提出古生物學家被少數的資料所混淆。他的警告當時被沒有被接受,但後來針對化石層的研究工作,證明早期的相關研究有誤。目前大部分古生物學家已普遍接受撞擊事件是白堊紀末滅絕事件的主要原因。不過,路易斯·阿爾瓦雷茨的兒子沃爾特·阿爾瓦雷茨認為在這次撞擊事件前,地球上已發生其他重大事件,例如:海平面下降、造成德干暗色岩的大規模火山爆發,它們可能也是這次滅絕事件的綜合原因之一[129]。
生態系統變遷
在2008年,Graeme T. Lloyd等人提出假設,認為開花植物的出現,造成白堊紀晚期的恐龍多樣性降低。在白堊紀早期以前,陸地的優勢植物是裸子植物,例如針葉樹;在白堊紀中晚期,開花植物開始成為優勢植物之一,但其地理範圍有限。根據恐龍的糞化石顯示,白堊紀晚期的大部分草食性恐龍仍以裸子植物為食,只有少部分以開花植物為食。同時,草食性昆蟲、哺乳動物的物種數量迅速增加,顯示牠們已經適應以這種新型態植物為食。Lloyd等人假設,相對於這些動物的多樣性增加,白堊紀晚期的恐龍多樣性則降低,這可能是導因於緩慢變遷的植物生態系統[48]。
古新世倖存者?
在K-T界線以上的地層中,偶爾會發現恐龍(不包含鳥類)的化石。在2001年,古生物學家J.E. Zielinski與J.R. Budahn宣稱在新墨西哥州聖胡安盆地發現一個鴨嘴龍科的股骨化石,並宣稱這是古新世恐龍的證據。該地層的年代為早古新世,接近6450萬年前。如果該骨頭不是因為風化作用而被重新沉積到該地層的話,它將被視為少數恐龍仍存活到新生代至少前50萬年的證據[130]。除此之外,在海爾河組也發現了其他恐龍化石,位在K-T界線以上1.3公尺,年代是K-T界線後約4萬年;世界其他各地也有類似發現,包含中國[131]。2004年的一项研究表明在中国南雄盆地发现的鸭嘴龙化石和蛋化石中发现有第三纪的花粉沉积。[132]
但是,許多科學家認為這些古新世恐龍的化石零散、不是較完整的骨骼,可能是被沖積作用帶離原本地點,然後再被重新掩埋[133][134]。
发掘历史
早在數千年前,人類就已發現恐龍的化石,但當時並不曉得這些石頭的真實面目。古代中國人將恐龍化石認為是中國神話中的龍骨頭,稱作“龍骨”,並在史書中留下相關紀錄。例如在東晉時期成書的《華陽國志》中,作者常璩便記錄到,四川武城曾經發現龍的骨頭[135]。中國人已挖掘恐龍化石多年,認為這些骨頭來自於龍,可當作中醫的藥材[136]。在歐洲,恐龍化石則被認為是神話中的巨人遺骸,或是大洪水時期前的大型動物。居住於古代中亞地區的西徐安人,可能將當地發現的原角龍化石,描述成獅子身體、大型爪、以及鷹頭,且守衛黃金的生物,成為獅鷲的形象來源[137]。

1677年的書籍《Natural History of Oxfordshire》封面。左邊是羅伯特·波爾蒂所繪的斑龍股骨末端,是最早被科學研究的恐龍化石。
在17世紀晚期的英格蘭,一些恐龍化石受到科學家的注意,開始初步的學術研究。在1676年,一個化石出土於英國牛津市附近的Cornwell的一處石灰岩採石場。這個骨頭碎片被交給牛津大學的化學教授羅伯特·波爾蒂(Robert Plot),他同時也是阿什莫爾博物館的館長,他在1677年的《Natural History of Oxfordshire》一書上敘述了對於這些骨頭的敘述。他將這塊骨頭正確地鑑定为一隻大型動物的股骨最下端,波爾蒂認為這些骨頭過大,所以並不屬於任何已知物種。波爾蒂後來認為這塊骨頭是個巨大人類的股骨,生存於聖經的大洪水事件之前[138]。在1763年,理查德·布魯克斯(Richard Brookes)再度敘述這個骨頭。他將這塊骨頭命名為Scrotum humanum,因為牠們看起來類似人類的一對睪丸。但是這個學名已經超過50年不曾被任何科學期刊使用,已成為一個無效的遺失名(Nomen oblitum)。這塊骨頭之後就遺失了,但已留下足夠的敘述,使後世科學家得以確認牠們屬於斑龍的股骨[139]。在1699年,Edward Lhuyd在牛津市近郊發現一顆牙齒化石,命名為Rutellum implicatum[140][141]。在2002年,科學家發現這顆牙齒屬於蜥腳下目恐龍,成為比斑龍還早被命名的恐龍。但是,這個學名的建立早於林奈氏分類系統的建立,而且已經超過50年不曾被任何科學期刊使用,而且是個根據牙齒而被建立的學名,因此本身也是個無效的遺失名[142]。

威廉·巴克蘭畫像
在1815年到1824年期間,牛津大學的地質學家威廉·巴克蘭,收集更多的斑龍化石,並在1824年建立斑龍屬(Megalosaurus),成為第一種被正式命名、敘述的恐龍有效屬[143][138]。第二種被命名、敘述的恐龍則是禽龍,是由英國動物學家吉迪恩·曼特爾在1822年所發現、研究,曼特爾當時認為這些化石與鬣蜥有類似處。在1825年,吉迪恩·曼特爾建立禽龍屬(Iguanodon)[144][145]。
歐洲與北美洲的科學家開始關注於這些巨大蜥蜴化石。在1842年,英國古生物學家理查·歐文認為斑龍、禽龍、林龍等物種具有許多明顯共同特徵,因此建立恐龍(Dinosauria)一詞,以包含這三個屬。在維多利亞女王的丈夫艾伯特親王的支持下,歐文在倫敦南肯辛頓建立了自然歷史博物館,以展出英國的恐龍與其他動物的化石。
在1858年,正式發現第一個北美洲的恐龍化石。早在1838年,紐澤西州哈登菲爾德鎮的一個泥灰坑挖出大型的骨頭,直到20年後,化石收藏家威廉·帕克·佛克(William Parker Foulke)在當地挖出更多的化石,並由古生物學家約瑟夫·萊迪命名為鴨嘴龍。這副鴨嘴龍標本是首次發現的接近完整恐龍化石,也顯示牠們是種二足動物,在此之前,大部分科學家認為恐龍是群類似蜥蜴的四足動物。佛克發現的鴨嘴龍標本,開啟了美國的恐龍狂潮。

奧塞內爾·查利斯·馬什,19世紀後期化石戰爭的兩位競爭者之一

愛德華·德林克·科普,化石戰爭的另一位競爭者
在19世紀後期的恐龍狂潮中,奧塞內爾·查利斯·馬什與愛德華·德林克·科普間有激烈的競爭,兩人競相發現新的恐龍,這段著名的競爭稱為化石戰爭。戰爭的導火線是馬什公開指出科普的薄板龍重建有嚴重的錯誤,科普將這隻蛇頸龍類的頭部錯置到尾巴末端。兩人的競爭長達30年,直到科普在1897年死亡,科普已將他的所有資產投注於恐龍的挖掘工作中,而馬什則得到美國地質調查局的協助,有較好的發現。但許多恐龍標本,因為兩人的粗略挖掘與處理步驟,而遭到損毀或破壞,例如採用火藥來挖掘化石。儘管馬什與科普的挖掘工作不完善,但他們的貢獻非常巨大,馬什命名了86個恐龍的新種,科普則命名了56個,共計142個新種恐龍。馬什發現的化石現正存放於耶魯大學的皮博迪自然史博物馆,科普的化石則存放在紐約美國自然歷史博物館[146]。
自1897年起,恐龍化石已在各大洲發現,甚至包含南極洲。第一個發現的南極州恐龍是甲龍科的南極甲龍,是在1986年發現於羅斯島;但在1994年,獸腳亞目的冰脊龍首先在科學期刊中被正式命名、敘述,成為第一個被命名的南極洲恐龍。自20世紀起,非洲和亞洲的挖掘工作開始展開。1907年,非洲的坦德古魯(位於坦尙尼亞境內)地區出土了巨大的骸骨。1908至1912年間,柏林自然史博物館的艾德文·亨尼希和韋爾納·亞南施來到這裡探勘,又挖掘出一隻腕龍屬的巨大恐龍,以及一隻劍龍屬的肯特龍骸骨。在亞洲,蒙古的戈壁沙漠裡,找到原角龍的蛋和窩。1922至1925年間,美國自然史博物館的工作隊在戈壁考察,又發現了其他種類的恐龍骸骨。[147]
目前的恐龍挖掘熱區有南美洲南部(尤其是阿根廷)與中國。中國發現了許多有羽毛恐龍標本,當時的乾燥氣候有助於保存化石與化石化的過程。
恐龍文藝復興
開始於60年代後期的恐龍文藝復興,對於恐龍的生理學研究產生重大的影響。這個科學革命始於約翰·奧斯特倫姆(John Ostrom)發現恐爪龍是種活躍的掠食動物,而且可能是種恆溫動物,與過去所認定的恐龍是種慵懶的變溫動物,有很大的不同。古脊椎動物學界開始熱烈爭論恐龍的相關學問,進而成為全球性的顯學。目前的主要新化石挖掘處,多是在過去沒被探索過的區域,例如:印度、南美洲、馬達加斯加、南極洲、尤其是中國,大部分的新發現有羽毛恐龍多出自於中國。親緣分支分類法是以不同生物的特徵來確定彼此關係,也成功地應用在恐龍的分類上。親緣分支分類法與其他現代科學方法,有助於解決不完整、破碎化石所產生的許多疑點。
大眾文化

一個19世紀的斑龍雕像,位於倫敦水晶宮公園
對於人類而言,恐龍是種充滿神秘感的巨大生物。因此恐龍已成為大眾文化與媒體的一部分。在1854年,當時為維多利亞時代,倫敦水晶宮公園就建立了數座恐龍雕像,將恐龍重建為類似蜥蜴的大型四足動物。這些雕像在一般大眾間造成轟動,當時也有販售小型模型,這是最早的周邊商品之一。自水晶宮公園之後,世界各地的公園與博物館多有恐龍展覽,這些展覽更使當時大眾增加對恐龍的興趣[148]。大眾對於恐龍的興趣有助於恐龍科學的發展。博物館之間的競爭,使馬什與科普間的化石戰爭激烈化,兩人極力發現更多、更奇特的化石,這有助於19世紀後期古生物學的發展[149]。從查爾斯·耐特(Charles R. Knight)的一系列恐龍繪畫可以看出,在19世紀後期,恐龍已被視為活躍的動物[150]。耐特的繪畫也可看出,當時古生物學界已將部份恐龍視為二足動物,採取尾巴拖曳在地表的三腳架步態[151]。
首次提到恐龍的文學作品是查尔斯·狄更斯在1852年出版的《荒凉山庄》,該小說的開場提到了斑龙[152]。恐龍還出現在許多奇幻小說中,著名的有:阿瑟·柯南·道尔在1912年出版的《失落的世界》、迈克尔·克莱顿在1990年出版的《侏羅紀公園》,兩者都曾被改編為電影。一些古生物學家出版的大眾恐龍讀物,則使一般大眾對恐龍、科學產生興趣,尤其是兒童[153]。
恐龍常是电影與動畫電影的主題之一。恐龍出現於電影中,最早可追溯到1914年的動畫電影《恐龍葛蒂》[154],以及同年的《蛮力》(Brute Force),後者更首次創造出恐龍與原始人類打鬥的場景[153]。在早期電影中,與恐龍有關的最著名電影分別為:1925年的《失落的世界》、以及1933年的《金剛》,後者因暴龍與金剛打鬥的場景而出名,並採用了模型、單格拍攝、接畫、背景拍攝…等多種特殊效果[153]。在1950年代到1960年代,恐龍多次成為怪獸電影的題材,例如:1953年的《原子怪獸》(The Beast from 20,000 Fathoms)、1956年的《荒山大賊王》(The Beast of Hollow Mountain)、1966年的《大洪荒》(One Million Years B.C.)、1969年的《暴龍關吉》(The Valley of Gwangi)、1975年的《被時間遺忘的土地》(The Land That Time Forgot)。自1954年開始的日本电影《哥斯拉》與其後續作品,其怪獸哥吉拉即是參考暴龍、禽龙、劍龍等恐龍創造出來,1998年上映的美國電影《酷斯拉》則是由哥吉拉所修改改編而成[155]。另外,恐龍也出現在1940年的迪士尼動畫電影《幻想曲》的「春之祭」段落[153]。
1993年的電影《侏儸紀公園》,是近年最具影響力的恐龍電影,曾位居最高電影票房榜首達四年之久,並在電腦繪圖技術有重大突破;《侏羅紀公園》與其系列作品將一些70年代以來的新假設、發現,透過電影傳播給一般大眾,例如:活躍的形象、二足恐龍與翼龍類的與正確步態、某些恐龍可能是高智商與群居的動物、比暴龍更大型的棘龙等[153][156]。近年以恐龍為主題的電影或動畫電影,另有《歷險小恐龍》、《恐龍》、《大雄的恐龍》、《大雄與龍之騎士》、《冰原歷險記3:恐龍現身》、《恐龍當家》。
拜先進的電腦繪圖技術與降低的成本所賜,近年有許多討論恐龍或史前生物的電視節目,例如:《与恐龙同行》、《恐龍紀元》、《恐龍兇面目》、《恐龍星球》、《史前公園》、《与巨兽同行》、《海底霸王》、以及《動物末日》。恐龍還出現在許多兒童節目或動畫中,例如:《摩登原始人》、《恐龍戰隊》、《爆龍戰隊》、《獸電戰隊》、《金剛戰士》、《巴尼与朋友》……。恐龍也成為眾多商品、玩具、電玩遊戲的主題,例如:《恐龙新世纪》、《恐龍危機》、《方舟:生存進化》。
衍生用語
- 恐龍妹:指相貌不出眾、體態肥胖的女性。
恐龍法官:指司法界中對現實反應有別於當代社會民情的法官。
宗教观点
许多宗教团体关于恐龙的观点与大多数科学家的共识不一样,这主要是由于科学界的看法与經文中的創造論故事相抵觸而造成的,伊卡黑石、阿坎巴羅雕像等歐帕茲是該些團體常用的假例證[157][158][159]。然而,科学界的主流不认同那些关于恐龙的宗教解释[160][161]。
相关条目
- 恐龙列表
参考文献
^ Borenstein, Seth. Study traces dinosaur evolution into early birds. AP News. 2014-07-31 [2015-03-08].
^ Lee, Michael S. Y.; Cau, Andrea; Naish, Darren; Dyke, Gareth J. Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds. Science. 1 August 2014, 345 (6196): 562–566 [2 August 2014]. doi:10.1126/science.1252243.
^ 『鳥類学辞典』 (2004)、805-806頁
^ (英文)Gauthier, Jacques; de Querioz, Kevin. Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name 'Aves'. (PDF). New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom (PDF)使用|format=需要含有|url=(帮助). Peabody Museum of Natural History, Yale University. [2001] [2009-09-22]. ISBN 0-912532-57-2.
^ 5.05.15.2 (英文)Feduccia, Alan. Birds are Dinosaurs: Simple Answer to a Complex Problem (PDF). The Auk (PDF)使用|format=需要含有|url=(帮助) 119(4). Department of Biology, University of North Carolina. : 1187–1201 [2002] [2009-09-22].
^ (英文)Bakker, R. T., Galton, P., 1974. Dinosaur monophyly and a new class of vertebrates. Nature 248:168-172.
^ (英文)Glut, Donald F. Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. 1997: 40. ISBN 0-89950-917-7.
^ (英文)Lambert, David; and the Diagram Group. The Dinosaur Data Book. New York: Avon Books. 1990: 288. ISBN 0-380-75896-2. 引文使用过时参数coauthors (帮助)
^ (英文)Morales, Michael. Nondinosaurian vertebrates of the Mesozoic. (编) Farlow, James O.; and Brett-Surman, Michael K. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. 1997: 607–624. ISBN 0-253-33349-0.
^ (英文)Hu Yaoming; Meng Jin; Wang Yuanqing; and Li Chuankui. Large Mesozoic mammals fed on dinosaurs. Nature. 2005, 433: 149–152. doi:10.1038/nature03102. 引文使用过时参数coauthors (帮助)
^ (英文)Wang, S.C., and Dodson, P. Estimating the Diversity of Dinosaurs. Proceedings of the National Academy of Sciences USA. 2006, 103 (37): 13601–13605. PMID 16954187.
^ (英文)Russell, Dale A. China and the lost worlds of the dinosaurian era. Historical Biology. 1995, 10: 3–12.
^ (英文)恐龍數量統計的爭議與困難?、BBC, September 17, 2008
^ 14.014.114.2 (英文)MacLeod, N, Rawson, PF, Forey, PL, Banner, FT, Boudagher-Fadel, MK, Bown, PR, Burnett, JA, Chambers, P, Culver, S, Evans, SE, Jeffery, C, Kaminski, MA, Lord, AR, Milner, AC, Milner, AR, Morris, N, Owen, E, Rosen, BR, Smith, AB, Taylor, PD, Urquhart, E & Young, JR. The Cretaceous–Tertiary biotic transition. Journal of the Geological Society. 1997, 154 (2): 265–292.
^ 關於恐龍十大有趣事實:有些恐龍能活到300歲 - 中時電子報
^ (英文)Owen, R. (1842). "Report on British Fossil Reptiles." Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
^ (英文)Liddell-Scott-Jones Lexicon of Classical Greek. [2008-08-05].
^ (英文)Farlow, J.O., and Brett-Surman, M.K. Preface. In: Farlow, J.O., and Brett-Surman, M.K. (eds.). The Complete Dinosaur.. Indiana University Press. 1997: ix–xi. ISBN 0-253-33349-0.
^ 甄朔南. 中國恐龍時代. 書泉. 2003. ISBN 986-12-1058-X.
^ 20.020.120.220.320.4 (英文)Benton, Michael J. Origin and relationships of Dinosauria. (编) Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria 2nd. Berkeley: University of California Press. 2004: 7–19. ISBN 0-520-24209-2.
^ 21.021.1 (英文)Holtz, Jr., T.R. Classification and evolution of the dinosaur groups. In: Paul, G.S. (ed.). The Scientific American Book of Dinosaurs.. St. Martin's Press. 2000: 140–168. ISBN 0-312-26226-4.
^ (英文)Langer, M.C., Abdala, F., Richter, M., and Benton, M.J. A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil. Comptes Rendus de l'Academie des Sciences, Paris: Sciences de la terre et des planètes. 1999, 329: 511–517.
^ (英文)Nesbitt, Sterling J.; Irmis, Randall B.; and Parker, William G. A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology. 2007, 5 (2): 209–243. 引文使用过时参数coauthors (帮助)
^ (英文)Dr. Holland and the Sprawling Sauropods. (原始内容存档于2011-06-12).
^ 25.025.1 (英文)Benton, M.J. Vertebrate Paleontology. Blackwell Publishers. 2004: xii–452. ISBN 0-632-05614-2.
^ (英文)Cowen, Richard. Dinosaurs. History of Life 4th edition. Blackwell Publishing. : 151–175. ISBN 1405117567. 引文格式1维护:冗余文本 (link)
^ 27.027.1 (英文)Kubo, T.; and Benton, M.J. Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates. Palaeontology. 2007, 50 (6): 1519–1529. 引文使用过时参数coauthors (帮助)
^ 28.028.1 (英文)Benton, Michael J. Origin and relationships of Dinosauria. (编) Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria 2nd. Berkeley: University of California Press. 2004: 7–19. ISBN 0-520-24209-2.
^ (英文)Olshevsky, G. (2000). "An annotated checklist of dinosaur species by continent." Mesozoic Meanderings, 3: 1–157
^ 30.030.130.230.330.430.530.6 (英文)Padian, K. (2004). Basal Avialae. In: Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria(second edition). University of California Press:Berkeley, 210–231. ISBN 978-0-520-24209-8.
^ (英文)Seeley, H.G. On the classification of the fossil animals commonly named Dinosauria. Proc R Soc London (Royal Society). 1887, 43: 165–171.
^ (英文)Romer, A.S. Osteology of the Reptiles. University of Chicago. 1956.
^ (英文)Ostrom, J.H., The evidence of endothermy in dinosaurs, (编) Thomas, R.D.K. and Olson, E.C., A cold look at the warm-blooded dinosaurs (PDF), Boulder, CO: American Association for the Advancement of Science: 82–105, 1980, (原始内容 (PDF)存档于2008-05-27)
^ (英文)Bakker, R. T., and Galton, P. Dinosaur monophyly and a new class of vertebrates. Nature. 1974, 248: 168–172. doi:10.1038/248168a0.
^ (英文)Citation for Permian/Triassic extinction event, percentage of animal species that went extinct. See commentary
^ (英文)Another citation for P/T event data. See commentary
^ 37.037.1 (英文)Sereno PC. The evolution of dinosaurs. Science. 1999, 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137.
^ (英文)Sereno, P.C.; Forster, C.A.; Rogers, R.R.; and Monetta, A.M. Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. Nature. 1993, 361: 64–66. doi:10.1038/361064a0. 引文使用过时参数coauthors (帮助)
^ (英文)Brusatte, S.L.; Benton, M.J.; Ruta, M.; and Lloyd, G.T. Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science. 2008, 321 (5895): 1485–1488. doi:10.1126/science.1161833. 引文使用过时参数coauthors (帮助)
^ (英文)Paul, G.S. Predatory Dinosaurs of the World. New York: Simon and Schuster. 1988: 248–250.
^ (英文)Clark, J.M., Maryanska, T., and Barsbold, R. (2004). "Therizinosauroidea", in The Dinosauria, 2nd ed. 151–164.
^ (英文)Norell, M.A., and Makovicky, P.J. (2004). "Dromaeosauridae", in The Dinosauria, 2nd ed. 196-210.
^ 43.043.143.243.3 (英文)Holtz, Thomas R., Jr.; Chapman, Ralph E.; and Lamanna, Matthew C. Mesozoic biogeography of Dinosauria. (编) Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria 2nd. Berkeley: University of California Press. 2004: 627–642. ISBN 0-520-24209-2. 引文使用过时参数coauthors (帮助)
^ 44.044.144.244.344.4 (英文)Fastovsky, David E.; and Smith, Joshua B. Dinosaur paleoecology. (编) Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria 2nd. Berkeley: University of California Press. 2004: 614–626. ISBN 0-520-24209-2. 引文使用过时参数coauthors (帮助)
^ (英文)Sereno, P.C.; Wilson, J.A.; Witmer, L.M.; Whitlock, J.A.; Maga, A.; Ide, O.; and Rowe, T.A. Structural extremes in a Cretaceous dinosaur. PLoS ONE. 2007, 2 (11): e1230. doi:10.1371/journal.pone.0001230. 引文使用过时参数coauthors (帮助)
^ (英文)Prasad, V.; Strömberg, C.A.E.; Alimohammadian, H; and Sahni, A. Dinosaur coprolites and the early evolution of grasses and grazers. Science. 2005, 310 (5751): 1170–1180. doi:10.1126/science.1118806. 引文使用过时参数coauthors (帮助)
^ (英文)Archibald, J. David; and Fastovsky, David E. Dinosaur Extinction. (编) Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria 2nd. Berkeley: University of California Press. 2004: 672–684. ISBN 0-520-24209-2. 引文使用过时参数coauthors (帮助)
^ 48.048.1 (英文)Lloyd, G.T., Davis, K.E., Pisani, D. Dinosaurs and the Cretaceous Terrestrial Revolution. Proceedings of the Royal Society: Biology. 22 July 2008, 275: 2483 [2008-07-28]. doi:10.1098/rspb.2008.0715. The URL contains links to all the content, and it's all free.
^ 49.049.1 (英文)Dal Sasso, C. and Signore, M. Exceptional soft-tissue preservation in a theropod dinosaur from Italy. Nature. 1998), 292 (6674): 383–387. doi:10.1038/32884. 请检查|date=中的日期值 (帮助)
^ 50.050.150.2 (英文)Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science. 2005, 307 (5717): 1952–1955. doi:10.1126/science.1108397.
^ (英文)Assessment and Update Status Report on the Blue Whale Balaenoptera musculus (PDF). Committee on the Status of Endangered Wildlife in Canada. 2002 [2007-12-05]. (原始内容 (PDF)存档于2004-05-21).
^ 52.052.152.2 (英文)Carpenter, Kenneth. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus. (编) Foster, John R.; and Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation (pdf). New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque: New Mexico Museum of Natural History and Science. 2006: 131–138. [失效連結]
^ (英文)Farlow, James A. On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs. (编) Dodson, Peter; and Gingerich, Philip. Functional Morphology and Evolution. American Journal of Science, Special Volume 293-A. 1993: 167–199.
^ (英文)Peczkis, J. (1994). "Implications of body-mass estimates for dinosaurs". Journal of Vertebrate Paleontology 14(4): 520-33
^ (英文)Anatomy and evolution. National Museum of Natural History. [2007-11-21].
^ Colbert, E.H. (1968). Men and Dinosaurs: The Search in Field and Laboratory. E. P. Dutton & Company:New York, vii + 283 p. ISBN 978-0-14-021288-4.
^ (英文)Lovelace, David M.; Hartman, Scott A.; and Wahl, William R. Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny. Arquivos do Museu Nacional. 2007, 65 (4): 527–544. 引文使用过时参数coauthors (帮助)
^ (英文)dal Sasso, C., Maganuco, S., Buffetaut, E., and Mendez, M.A. (2006). New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities. Journal of Vertebrate Paleontology 25(4):888–896.
^ 59.059.1 (英文)Zhang, Fucheng; Zhou, Zhonghe; Xu, Xing; Wang, Xiaolin and Sullivan, Corwin. "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers". <http://www.nature.com/nature/journal/v455/n7216/full/nature07447.html> Nature 455, 1105-1108 (23 October 2008) | doi:10.1038/nature07447
^ 60.060.1 (英文)Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. (2009). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin." Chinese Science Bulletin, six pages, accepted November 15, 2008.
^ (英文)Holtz, Thomas R. Jr. (2008) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages Supplementary Information
^ (英文)Butler, R.J. and Zhao, Q. "The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China." Cretaceous Research, Corrected Proof, Available online 8 April 2008 doi:10.1016/j.cretres.2008.03.002
^ (法文)Yans J, Dejax J, Pons D, Dupuis C & Taquet P. Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique). Comptes Rendus Palevol. 2005, 4 (1-2): 135–150. doi:10.1016/j.crpv.2004.12.003.
^ (英文)Day, J.J.; and Upchurch, P. Sauropod trackways, evolution, and behavior. Science. 2002, 296: 1659. doi:10.1126/science.107016. 引文使用过时参数coauthors (帮助)
^ (英文)Wright, Joanna L. Steps in understanding sauropod biology. (编) Curry Rogers, Kristina A.; and Wilson, Jeffrey A. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. 2005: 252–284. ISBN 0-520-24623-3.
^ (英文)Varricchio, D.J.; Sereno, P.C.; X. Zhao; L. Tan; Wilson, J.A.; and Lyon, G.H. Mud-trapped herd captures evidence of distinctive dinosaur sociality (pdf). Acta Palaeontologica Polonica. 2008, 53 (4): 567–578 [2008-12-09]. 引文使用过时参数coauthors (帮助)
^ (英文)Lessem, Don; and Glut, Donald F. Allosaurus. The Dinosaur Society's Dinosaur Encyclopedia. Random House. 1993: 19–20. ISBN 0-679-41770-2. 引文使用过时参数coauthors (帮助)
^ (英文)Maxwell, W. D.; Ostrom, J.H. Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. Journal of Vertebrate Paleontology. 1995, 15 (4): 707–712. 引文使用过时参数coauthors (帮助)(abstract 互联网档案馆的存檔,存档日期2007-09-27.)
^ (英文)Roach, Brian T.; and Brinkman, Daniel L. A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. Bulletin of the Peabody Museum of Natural History. 2007, 48 (1): 103–138. 引文使用过时参数coauthors (帮助)
^ (英文)Horner, J.R.; and Makela, R. Nest of juveniles provides evidence of family-structure among dinosaurs. Nature. 1979, 282 (5736): 296–298. 引文使用过时参数coauthors (帮助)
^ (英文)Chiappe, Luis M.; Jackson, Frankie; Coria, Rodolfo A.; and Dingus, Lowell. Nesting titanosaurs from Auca Mahuevo and adjacent sites. (编) Curry Rogers, Kristina A.; and Wilson, Jeffrey A. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. 2005: 285–302. ISBN 0-520-24623-3. 引文使用过时参数coauthors (帮助)
^ (英文)偷蛋龍或原角龍的蛋巢?
^ (英文)Meng Qingjin; Liu Jinyuan; Varricchio, David J.; Huang, Timothy; and Gao Chunling. Parental care in an ornithischian dinosaur. Nature. 2004, 431: 145–146.
^ (英文)Reisz RR, Scott, D Sues, H-D, Evans, DC & Raath, MA. Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance. Science. 2005, 309: 761–764. doi:10.1126/science.1114942. 使用|accessdate=需要含有|url=(帮助)
^ (英文)遷徙中的恐龍家庭
^ (英文)Tanke, Darren H.; and Currie, Philip J. Head-biting behavior in theropod dinosaurs: paleopathological evidence (PDF). Gaia. 1998, (15): 167–184. ISSN 0871-5424. (原始内容 (pdf)存档于2008-02-27). 引文使用过时参数coauthors (帮助)
^ (英文)Hopson, James A. The evolution of cranial display structures in hadrosaurian dinosaurs. Paleobiology. 1975, 1 (1): 21–43.
^ (英文)Diegert, Carl F.; and Williamson, Thomas E. A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen. Journal of Vertebrate Paleontology. 1998, 18 (3, Suppl.): 38A. 引文使用过时参数coauthors (帮助)
^ (英文)搏鬥中的恐龍. American Museum of Natural History. [2007-12-05]. (原始内容存档于2012-01-18).
^ 80.080.1 (英文)Carpenter, K. Evidence of predatory behavior by theropod dinosaurs.. Gaia. 1998, 15: 135–144 [2007-12-05]. (原始内容存档于2007-11-17).
^ (英文)Rogers, Raymond R.; Krause, David W.; & Curry Rogers, Kristina. Cannibalism in the Madagascan dinosaur Majungatholus atopus. Nature. 2007, 422 (6931): 515–518. doi:10.1038/nature01532. 引文使用过时参数coauthors (帮助)
^ (英文)Varricchio DJ, Martin, AJ and Katsura, Y. First trace and body fossil evidence of a burrowing, denning dinosaur. Proceedings of the Royal Society B: Biological Sciences. 2007, 274 (1616): 1361–1368. doi:10.1098/rspb.2006.0443.
^ (英文)Chatterjee, S.; and Templin, R.J. Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui (pdf). Proceedings of the National Academy of Sciences. 2007, 104 (5): 1576–1580. 引文使用过时参数coauthors (帮助)
^ (英文)Zhang, F.; Zhou, Z.; Xu, X.; and Wang, X. A juvenile coelurosaurian theropod from China indicates arboreal habits. Naturwissenschaften. 2002, 89 (9): 394–398. doi:10.1007/s00114-002-0353-8.
^ (英文)Alexander RM. Dinosaur biomechanics. Proceedings of the Royal Society of Biological Sciences. 2006, 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532.
^ (英文)Goriely A & McMillen T. Shape of a cracking whip. Physical Review Letters. 2002, 88 (24): 244301. doi:10.1103/PhysRevLett.88.244301.
^ (英文)Henderson, D.M. Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis.. Canadian Journal of Zoology. 2003, 81 (8): 1346–1357. doi:10.1139/z03-122.
^ (英文)Parsons, K.M. (2001). Drawing Out Leviathan. Indiana University Press. 22–48. ISBN 978-0-253-33937-9.
^ (英文)Fisher, P. E., Russell, D. A., Stoskopf, M. K., Barrick, R. E., Hammer, M. & Kuzmitz, A. A. Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. Science. 2000, 288 (5465): 503–505. doi:10.1126/science.288.5465.503.
^ (英文)Hillenius, W. J. & Ruben, J. A. The evolution of endothermy in terrestrial vertebrates: Who? when? why?. Physiological and Biochemical Zoology. 2004, 77 (6): 1019–1042. doi:10.1086/425185.
^ (英文)Rowe T, McBride EF, & Sereno PC. Dinosaur with a Heart of Stone. Science. 2001, 291 (5505): 783. doi:10.1126/science.291.5505.783a.
^ (英文)Chinsamy, Anusuya; and Hillenius, Willem J. (2004). "Physiology of nonavian dinosaurs". The Dinosauria, 2nd. 643–659.
^ (英文)MKaye TG, Gaugler G, Sawlowicz Z. Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms.. PLoS ONE. July 2008, 3 (7): e2808 [2008-07-29]. doi:10.1371/journal.pone.0002808.
^ (英文)新研究衝擊現存恐龍軟組織的說法Newswise, Retrieved on 29 July 2008.
^ (英文)Wang, H., Yan, Z. and Jin, D. Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil. Molecular Biology and Evolution. 1997, 14: 589–591 [2007-12-05]. (原始内容存档于2008-08-28).
^ (英文)Chang BS, Jönsson K, Kazmi MA, Donoghue MJ, Sakmar TP. Recreating a Functional Ancestral Archosaur Visual Pigment. Molecular Biology and Evolution. 2002, 19 (9): 1483–1489 [2007-12-05]. PMID 12200476.
^ (英文)Embery G, Milner AC, Waddington RJ, Hall RC, Langley MS, Milan AM. Identification of proteinaceous material in the bone of the dinosaur Iguanodon. Connect Tissue Res. 2003, 44 (Suppl 1): 41–6. PMID 12952172.
^ (英文)Schweitzer MH, Marshall M, Carron K, Bohle DS, Busse SC, Arnold EV, Barnard D, Horner JR, Starkey JR. Heme compounds in dinosaur trabecular bone. Proc Natl Acad Sci U S A. 1997, 94 (12): 6291–6. PMID 9177210.
^ (英文)Huxley, Thomas H. On the animals which are most nearly intermediate between birds and reptiles. Annals of the Magazine of Natural History. 1868, 4 (2): 66–75.
^ (英文)Heilmann, Gerhard. The Origin of Birds. London: Witherby. 1926: 208pp.
^ (英文)Osborn, Henry Fairfield. Three new Theropoda, Protoceratops zone, central Mongolia (pdf). American Museum Novitates. 1924, 144: 1–12.
^ (英文)Ostrom, John H. The ancestry of birds. Nature. 1973, 242 (5393): 136. doi:10.1038/242136a0.
^ (英文)Gauthier, Jacques. Saurischian monophyly and the origin of birds. (编) Padian, Kevin. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. 1986: 1–55.
^ (英文)Mayr, G., Pohl, B. and Peters, D.S. A Well-Preserved Archaeopteryx Specimen with Theropod Features. Science. 2005, 310 (5753): 1483–1486. doi:10.1126/science.1120331.
^ (英文)Martin, Larry D. A basal archosaurian origin for birds. Acta Zoologica Sinica. 2006, 50 (6): 977–990.
^ (英文)Wellnhofer, P. Ein neuer Exemplar von Archaeopteryx. Archaeopteryx. 1988, 6: 1–30.
^ (英文)Xu X.; Norell, M.A.; Kuang X.; Wang X.; Zhao Q.; and Jia C. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature. 2004, 431 (7009): 680–684. PMID 15470426.
^ (英文)Göhlich, U.B.; and Chiappe, L.M. A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago. Nature. 2006, 440: 329–332. 引文使用过时参数coauthors (帮助)
^ (英文)Lingham-Soliar, T. The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers. Naturwissenschaften. 2003, 90 (12): 563–567. PMID 14676953.
^ 110.0110.1 (英文)Feduccia, A.; Lingham-Soliar, T.; and Hinchliffe, J.R. Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. Journal of Morphology. 2005, 266 (2): 125–166. PMID 16217748. 引文使用过时参数coauthors (帮助)
^ (英文)Lingham-Soliar, T.; Feduccia, A.; and Wang X. A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres. Proceedings of the Biological Sciences. 2007, 274 (1620): 1823–9. PMID 17521978. 引文使用过时参数coauthors (帮助)
^ (英文)Prum, Richard O. Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002. The Auk. April 2003, 120 (2): 550–61.
^ (英文)O'Connor PM & Claessens LPAM. Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. Nature. 2005, 436: 253–256. doi:10.1038/nature03716.
^ (英文)Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA 等. Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina. PLoS ONE. 2008, 3 (9): e3303 [2008-09-29]. doi:10.1371/journal.pone.0003303. 已忽略未知参数|month=(建议使用|date=) (帮助) 引文格式1维护:显式使用等标签 (link)
^ (英文)Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System Newswise, Retrieved on September 29, 2008.
^ (英文)Wings O. A review of gastrolith function with implications for fossil vertebrates and a revised classification (PDF). Palaeontologica Polonica. 2007, 52 (1): 1–16 [2007-11-24]. (原始内容 (PDF)存档于2008-03-07).
^ (英文)Schweitzer, M.H.; Wittmeyer, J.L.; and Horner, J.R. Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. Science. 2005, 308: 1456–1460. doi:10.1126/science.1112158. 引文使用过时参数coauthors (帮助)
^ (英文)Lee, Andrew H.; and Werning, Sarah. Sexual maturity in growing dinosaurs does not fit reptilian growth models. Proceedings of the National Academy of Sciences. 2008, 105 (2): 582–587. doi:10.1073/pnas.0708903105. 引文使用过时参数coauthors (帮助)
^ (英文)Xu, X. and Norell, M.A. A new troodontid dinosaur from China with avian-like sleeping posture. Nature. 2004, 431: 838–841. doi:10.1038/nature02898.
^ (英文)Miller KG, Kominz MA, Browning JV, Wright JD, Mountain GS, Katz ME, Sugarman PJ, Cramer BS, Christie-Blick N, Pekar SF. The Phanerozoic record of global sea-level change. Science. 2005, 310 (5752): 1293–8. PMID 16311326. doi:10.1126/science.1116412.
^ (英文)McArthura JM, Janssenb NMM, Rebouletc S, Lengd MJ, Thirlwalle MF & van de Shootbruggef B. Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian). Palaeogeography, Palaeoclimatology, Palaeoecology. 2007, 248 (3-4): 391–430. doi:10.1016/j.palaeo.2006.12.015.
^ (英文)Alvarez, LW, Alvarez, W, Asaro, F, and Michel, HV. Extraterrestrial cause for the Cretaceous–Tertiary extinction. Science. 1980, 208 (4448): 1095–1108. doi:10.1126/science.208.4448.1095.
^ (英文)Hildebrand, Alan R.; Penfield, Glen T.; Kring, David A.; Pilkington, Mark; Zanoguera, Antonio Camargo; Jacobsen, Stein B.; Boynton, William V. Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico. Geology. September 1991, 19 (9): 867–871.
^ (英文)Pope KO, Ocampo AC, Kinsland GL, Smith R. Surface expression of the Chicxulub crater. Geology. 1996, 24 (6): 527–30. PMID 11539331. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2.
^ (英文) P, Claeys; Goderis, S. Solar System: Lethal billiards. Nature. 2007-09-05, 449: 30–31. doi:10.1038/449030a. 引文使用过时参数coauthors (帮助); 使用|accessdate=需要含有|url=(帮助)
^ 126.0126.1 (英文)Koeberl, C & MacLeodKG. Catastrophic Events and Mass Extinctions. Geological Society of America. 2002. ISBN 0-8137-2356-6.
^ 127.0127.1 (英文)Hofman, C, Féraud, G & Courtillot, V. 40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps. Earth and Planetary Science Letters. 2000, 180: 13–27. doi:10.1016/S0012-821X(00)00159-X.
^ 128.0128.1128.2 (英文)Duncan, RA & Pyle, DG. Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary. Nature. 1988, 333: 841–843. doi:10.1038/333841a0.
^ (英文)Alvarez, W. T. rex and the Crater of Doom. Princeton University Press. 1997: 130–146. ISBN 978-0691016306.
^ (英文)Fassett, JE, Lucas, SG, Zielinski, RA, and Budahn, JR. Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA (PDF). Catastrophic events and mass extinctions, Lunar and Planetary Contribution. 2001, 1053: 45–46 [2007-05-18].
^ (英文)Sloan, R. E., Rigby, K,. Van Valen, L. M., Gabriel, Diane. Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek formation. Science. 1986, 232 (4750): 629–633 [2007-05-18]. doi:10.1126/science.232.4750.629.
^ (英文)BUCK Brenda J. ; HANSON Andrew D. ; HENGST Richard A. ; SHU-SHENG Hu ;. "Tertiary dinosaurs" in the Nanxiong Basin, Southern China, are reworked from the Cretaceous. The Journal of geology. 2004, 112 (1): 111–118. doi:10.1086/379695.
^ (英文)Fastovsky, David E.; and Sheehan, Peter M. Reply to comment on "The Extinction of the dinosaurs in North America" (pdf). GSA Today. 2005, 15: 11. 引文使用过时参数coauthors (帮助)
^ (英文)Sullivan, RM. No Paleocene dinosaurs in the San Juan Basin, New Mexico. Geological Society of America Abstracts with Programs. 2003, 35 (5): 15 [2007-07-02].
^ (中文)董枝明. Dinosaurian Faunas of China. 北京,海洋出版社. 1992. ISBN 978-3-540-52084-9.
^ (英文)恐龍骨頭被當作醫藥使用. BBC News. 2007 [2007-07-06].
^ (英文)Mayor, Adrienne. The First Fossil Hunters: Paleontology in Greek and Roman Times. Princeton: Princeton University Press. 2000. ISBN 0-691-05863-6.
^ 138.0138.1 (英文)Sarjeant, William A.S. The earliert discoveries. (编) Farlow, James O.; and Brett-Surman, Michael K. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. 1997: 3–11. ISBN 0-253-33349-0.
^ (英文)Halstead, L.B. (1970). "Scrotum humanum Brookes 1763 - the first named dinosaur." Journal of Insignificant Research, 5: 14-15.
^ (英文)Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann:London.
^ (英文)Gunther, R.T. (1945). Early Science in Oxford: Life and Letters of Edward Lhuyd, volume 14. Author:Oxford.
^ (英文)Delair, J.B., and Sarjeant, W.A.S. (2002). The earliest discoveries of dinosaurs: the records re-examined. Proceedings of the Geologists' Association 113:185-197.
^ (英文)Buckland, W. (1824). "Notice on the Megalosaurus or great Fossil Lizard of Stonesfield." Transactions of the Geological Society of London, series 2, vol. 1: 390–396.
^ (英文)Mantell, Gideon A. Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex.. Philosophical Transactions of the Royal Society. 1825, 115: 179–186 [2007-02-21]. doi:10.1098/rstl.1825.0010.
^ (英文)Sues, Hans-Dieter. European Dinosaur Hunters. (编) James Orville Farlow and M. K. Brett-Surman (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. 1997: 14. ISBN 0-253-33349-0.
^ (英文)Holmes T. Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science. Silver Burdett Press. 1996. ISBN 978-0382391477.
^ Michard, Jean-Guy. 第二章:尋找名分. 《恐龍:失去的侏羅紀王國》. 發現之旅. 20. 萬美君/譯 2002年2月5日初版六刷. 臺北: 時報文化. 1995年3月10日: 第39–40頁. ISBN 978-957-13-1582-9 (中文(台灣)).
^ (英文)Torrens, H.S. The dinosaurs and dinomania over 150 years. Modern Geology. 1993, 18 (2): 257–286.
^ (英文)Breithaupt, Brent H. First golden period in the USA." In: Currie, Philip J. & Padian, Kevin (Eds.). The Encyclopedia of Dinosaurs. University of California Press. 1997: 347–350. ISBN 978-0122268106.
^ (英文)Paul, Gregory S. The Art of Charles R. Knight. (编) Paul, Gregory S. (ed.). The Scientific American Book of Dinosaurs. New York: St. Martin's Press. 2000: 113–118. ISBN 0-312-26226-4.
^ (英文)Breithaupt, Brent H.; Chure, Daniel J.; and Southwell, Elizabeth H. AMNH 5753: the world's first free-standing theropod skeleton. Journal of Vertebrate Paleontology. 1999, 19 (3, Suppl.): p. 33A. 引文使用过时参数coauthors (帮助) 引文格式1维护:冗余文本 (link)
^ (英文)"London. Michaelmas term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborne Hill." From page 1 of Dickens, Charles J.H. (1852). Bleak House. London: Bradbury & Evans.
^ 153.0153.1153.2153.3153.4 (英文)Glut, Donald F.; Brett-Surman, Michael K. Dinosaurs and the media. The Complete Dinosaur. Bloomington and Indianapolis: Indiana University Press. 1997: 675–706. ISBN 0-253-33349-0. 引文使用过时参数coauthors (帮助)
^ (英文)Sarjeant, William A.S. Dinosaurs in fiction. (编) Tanke, Darren H.; and Carpenter, K. (eds.). Mesozoic Vertebrate Life. Bloomington and Indianapolis: Indiana University Press. 2001: 504–529. ISBN 0-253-33907-3.
^ (英文)Snider, Mike. 哥吉拉造成核能恐懼. USA Today (Gannett Corporation). 2006-08-29 [2007-02-21].
^ (英文)Lucas, Spencer G. Dinosaurs in the public eye. Dinosaurs: The Textbook 3rd. Boston: McGraw-Hill. 2000: 247–260. ISBN 0-07-303642-0.
^ (中文)「人類與恐龍曾同時代存在」的化石 原題:墨西哥2012之五 進化論 曠世奇謊,大紀元新聞網,2011年10月30日
^ (中文)史前巨人文明存在的可能性,大紀元新聞網,2004年5月17日
^ (中文)【新紀元】張海連:恐龍滅絕和超功能,大紀元新聞網,2010年4月2日
^ (英文)Kitcher, Philip. Abusing Science: The Case Against Creationism. MIT Press. 1983: 213. ISBN 978-0-262-61037-7.
^ (英文)Dawkins, Richard. The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe Without Design. W.W. Norton. 1996: 400. ISBN 978-0393315707.
延伸閱讀
(英文) Kevin Padian, and Philip J. Currie. (1997年)《Encyclopedia of Dinosaurs》Academic Press. ISBN 978-0-12-226810-6. (由專家撰寫).
(英文) Paul, Gregory S. (2000年)《The Scientific American Book of Dinosaurs'》St. Martin's Press. ISBN 978-0-312-26226-6.
(英文) Paul, Gregory S. (2002年)《Dinosaurs of the Air: The Evolution and Loss of flight in Dinosaurs and Birds》Baltimore: The Johns Hopkins University Press. ISBN 978-0-8018-6763-7.
(英文) Weishampel, David B. (2004年)《The Dinosauria》University of California Press; 2nd edition. ISBN 978-0-520-24209-8.
(繁体中文) Michard, Jean-Guy. 《恐龍:失去的侏羅紀王國》. 發現之旅. 20. 萬美君/譯. 臺北: 時報文化. 1995年. ISBN 978-957-13-1582-9.
外部链接
维基物种中的分类信息:恐龙 |
维基共享资源中相关的多媒体资源:恐龙 |
- 圖像
- 紀錄片-恐龍獵人(國語版)
The Art Gallery - 超過2000幅恐龍的插畫
Skeletal Drawing - 恐龍的骨骼重建、電腦繪畫
The Science and Art of Gregory S. Paul - 葛瑞格利·保恐龍與插畫網站
- 中文
- 化石网
- 中國恐龍網
- 延陵恐龍志
- 化石網 - 恐龍網
- 化石网>>化石网报道——古生物新闻中心
- 化石网>>中国恐龙
- 化石网论坛>>恐龙专区
- 英文
Enchanted Learning網站 - 恐龍的簡介、生理特徵、分類、各屬、問與答
恐龍與其他的史前生物 - 大英自然史博物館網站- 世界各地的恐龍新聞
恐龍的簡介 - 加州大學柏克萊分校古生物博物館網站- 美國生活科學網站
- Dino Russ's Lair網站
- 恐龍的新聞、搜尋資料庫、圖片、討論區
DinoData[失效連結] - 恐龍的簡介、分類、與特徵
Dinosauria On-Line恐龍的簡介、發音(依英文名稱排列)- 恐龍的圖片、繪畫、分類
Thescelosaurus! - 恐龍各主要分支的介紹與演化樹、圖片
Palaeos - 恐龍形態類(Dinosauromorpha)、恐龍的特徵與演化樹
The Dinosaur Encyclopaedia - 恐龍各屬的簡介- 與恐龍共舞官方網站
IMDb電影資料庫 - 有關恐龍的電影、動畫、電視節目
EvoWiki - 恐龍的特徵- 2008年版的恐龍列表
- Palaeontologia Electronica
| ||||||||||||||||||||||||||||||||||||||||||||||||||||

|


